|
This
tribe was proposed by Bendib & Minet (1999) to embrace a group of taxa that
had larvae without verrucae, this character state being interpreted as
apomorphic, given the prevalence of verrucae in the larvae of Arctiidae as a
whole and in the sister-family, the Lymantriidae. Other genera included by
Bendib & Minet where larvae were unknown, were associated by having
synapomorphies with genera with known larvae. These adult characters included an
adult resting posture with antennae exposed, not directed backwards, antennae
with a light-coloured subapical section, a forewing with a red discocellular
mark, a hindwing with Rs and M1 fused, a metascutum with membranous areas, an
abdomen in the male with lateral tufts of hairlike androconia, and a
corethrogyne in the female.
The
Bornean fauna has one of the taxa listed by Bendib & Minet, Aemene
Walker, revived from synonymy with Siccia
Walker but including Hyposiccia Hampson
and Parasiccia Hampson, the only two
other Old World taxa included by Bendib & Minet. Siccia and Aemene have a
corethrogyne in the female and are therefore placed within the subtribe
Clemensiiti.
Two
other groups of genera may belong in the Cisthenini, but it is not clear whether
their larvae lack verrucae. In the Garudinia
Moore generic complex, the two larval descriptions in Piepers & Snellen
(1904) are ambiguous. That for Pseudoblabes
Zeller suggests setae are sparse, but that for Byrsia Walker refers to a dense covering. No larval descriptions
have been located for the Macaduma Walker
group.
The
feature that is shared by all three Old World groups is the presence of
enlarged, well defined paratergal sclerites at the junction of the vinculum and
tegumen on each side, convex, rather lobed towards the exterior. It often
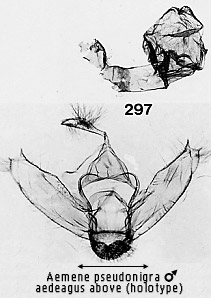
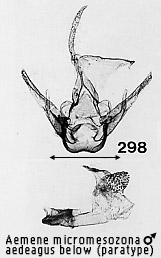
appears as if the vinculum is twisted into this expanded junction. The feature
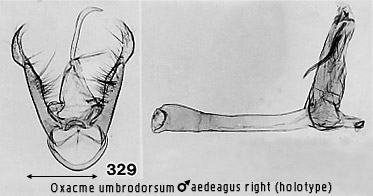
can be seen clearly in Figs 297, 298, 308, 315 and 329. It is not clear whether
this modification is homologous with the condition referred to by Bendib &
Minet for the Phryganopterygini, but they state it also occurs in Miltochrista
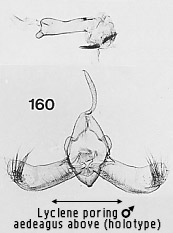
in the Nudariini. Amongst Nudariini studied for this work, it is rarely as
strongly developed as in the three Cisthenini groups just mentioned and always
appears less �lobed�. An example of strong development in the Nudariini is Lyclene poring sp. n. (Fig
160).
 |
 |
|
|
A
corethrogyne is widespread amongst females examined by Bendib & Minet
(1999), and is of distinctive form in the Garudinia
complex as discussed below. However, a corethrogyne can also occur in the
Nudariini, e.g. in two species groups of Lyclene
(see Lyclene biseriata
Hampson,
Lyclene apiseriata
sp.n.) where it may be homoplasious. It is present but only moderate in
the Macaduma group.
In
the Garudinia and Macaduma
groups Rs and M1 in the hindwing are more frequently stalked than
coincident, and the other potential apomorphies for groups within the tribe
listed by Bendib & Minet need further investigation, e.g. members of the Garudinia
complex do not have reddish discocellular marks on the forewing, and the
antennae of both groups lack subapical pale zones. The antennal resting posture
needs observation in the field.
The
Garudinia complex is defined by a
strong suite of characters in the male and female abdomen, many of which were
noted for the Scaptesyle generic
quartet discussed by Holloway (1984b). In the male there are slender apodemes on
the eighth tergite, sometimes separate, sometimes fused into a �V� or
�Y�. In the male genitalia the valves are deeply bifid, with a rather narrow
membranous dorsal part and a more strongly sclerotised, often spine-like
saccular part; there is a distinctive setose pouch-like structure between the
saccular bases (a possibly homologous structure is seen in Aemene pseudonigra sp. n. and �Utriculofera� macroplaga Hampson). The ductus ejaculatorius is
strongly sclerotised, a feature shared with the Macaduma group; sclerotisaton is moderate in some Aemene
and also some Nudariini. The female has a short ductus and extensive, but
irregular spining in the bursa (though similar features occur in the Nudariini),
but is particularly distinguished by the corethrogyne which has distinct zones
of straight and crinkled hairs (Fig 149).
Within
this Garudinia complex, many Oriental
genera and the widespread Padenia Moore
have a deep, often bilobed androconial hindwing in the male. The Scaptesyle
quartet is defined on its distinctive facies, the forewing having a broad
black border within which is a red or orange zone. Several Australasian genera
belong to the Garudinia group, most
endemic to, or more diverse in, New Guinea: Chiriphe
Walker; Padenodes Rothschild; Paradohertya
Bethune-Baker; Parascaptia Bethune-Baker;
Stenoscaptia Hampson.
The
Macaduma group (including,
tentatively, Utriculofera Hampson) has
a variably irregular forewing shape with all veins present except in Macaduma
where a radial sector vein has been lost (Fig 1f). In the other genera, R1
and R2 are independent, with (R3 (R4, R5)) branching. Macaduma
would fit this pattern if (R4, R5) were fused or one lost. The male abdomen
in all the genera has coremata, but these differ and are diagnostic in each
genus. The ductus ejaculatorius in the vesica is strongly sclerotised as in the Garudinia
group. There may be a moderate corethrogyne in many members of the group.
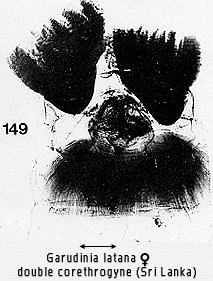 |
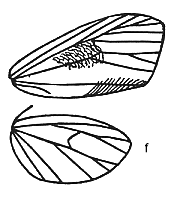
Fig 1f: Mantala
tineoides Walker |
>>Forward
<<Return to Contents page
|