|
The classification of the
Nolinae has been discussed by Holloway & Miller (1995), Holloway (1998) and
Holloway et al. (2001).
Most of the species are small, the forewings variegated black, grey and white,
often with areas of raised scales; more colourful species are seen in a few
genera such as Evonima
Walker. The male antennae are filiform and ciliate or bipectinate, the
former probably being the plesiomorphic state, and possibly secondarily derived
in Evonima. Those of the female are filiform except in
Dialithoptera Hampson where they are strongly bipectinate in both
sexes. Ocelli are weak or absent, strongest in
Beana
Walker, possibly the sister-genus to the rest of the subfamily.
The forewing venation usually lacks an areole except in Barasa
Walker and allies, though a small one may occur in more advanced taxa such as
some Meganola
Dyar. The radial sector veins posterior to R1 may be reduced to three or two
from four, and this was used by Hampson (1900) as the basis for his
classification. Reduction from three to two occurs within the more strict
definition of Nola
Leach of Holloway & Miller (1995). The hindwing is of the Nolidae groundplan
type in more basal genera, but with M3 and CuA1 stalked in all except
Beana,
and M2 separated from this. A trifine condition arises in more advanced genera
(mostly containing smaller species), probably through fusion of the stalked
veins as there is still a separate vein in the M2 position.
The male abdomen lacks tymbal organs but usually has apodemes on the eighth
tergite, and these may also occur on the sternite in more derived genera where
both sclerites can become distinctly narrowed. The genitalia have a simple uncus
and simple valves with a characteristically �noctuid� saccular harpe. The
saccular shield is often small, modified or lost, particularly where the
saccular bases meet broadly in the centre of the diaphragma. The most
characteristic feature seen in many genera is a pair of setose lobes that flank
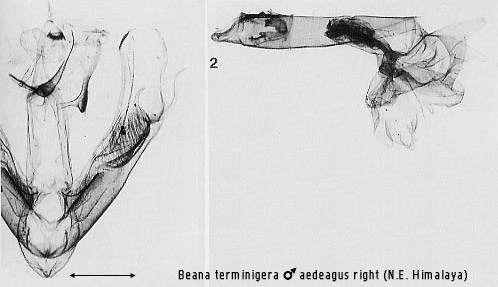
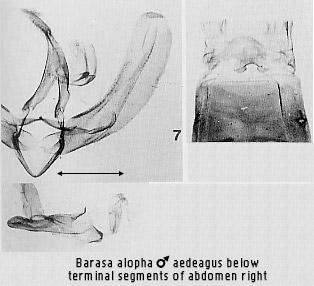
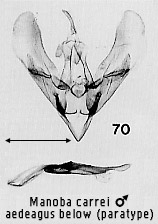
the scaphial area of the anal tube (e.g. Figs 2, 7, 70). Extreme modification of
the uncus is seen in Beana and
Ctenane
Swinhoe where it gives rise to rake-like structures, though these are located
differently and possibly not homologous. The uncus is lost in most
Nola,
which also have the valve diagnostically cleft. The aedeagus can have apical
projections, and the vesica may contain cornuti.
The female genitalia have no particularly diagnostic features in segments 8-10,
except the ovipositor lobes are usually short, quadrate or, seen together,
rather conical. The ductus and neck of the bursa are often slender, elongate.
There may be a small appendix bursae. The corpus bursae itself is usually
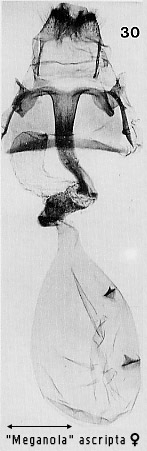
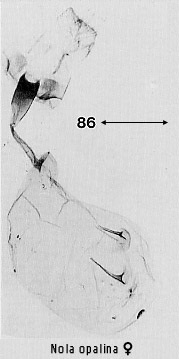
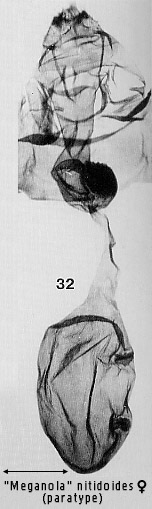
pyriform and contains signa, usually two, opposed scobinate corrugations (Fig 6)
or bands in more primitive genera, and invaginated ridges or thorn-like
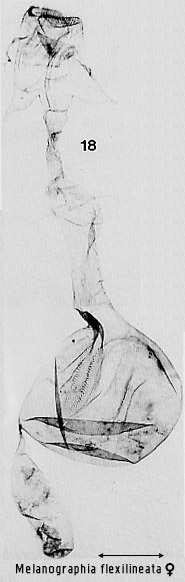
structures in more advanced ones (e.g. Figs 30, 86). Dialithoptera
is unusual in having two stellate signa similar to those of ennomine geometrids
(Fig 18).
The larva is characterised by a reduced anterior pair of abdominal prolegs and,
in all known larvae except that of Beana, secondary setae on verrucae. An
unusual feature recorded for a number of genera and species is the retention of
a stack of old larval head capsules in a horn on the �current� head (McFarland,
1980; Holloway & Miller, 1995; Holloway, 1998). This feature is recorded amongst
more derived members of the quadrifine sequence but is not known from trifine
genera except Evonima
and one example in Manoba.
The generic classification within the subfamily is far from satisfactory
(Holloway & Miller, 1995), with both nomenclatural confusion and a lack of a
clear understanding of morphological features. Genus-group names such as Nola,
Celama
Walker and Roeselia
H�bner have been applied indiscriminately in the past (e.g. Hampson, 1900; van
Eecke, 1926), with other genus-group names assigned as synonyms in a similar
fashion; Celama and Roeselia in fact fall within a strict
definition of Nola, leaving a considerable number of both quadrifine and
trifine genera outside this definition (old, erroneous concepts of Roeselia
and Celama respectively). The quadrifines were placed in Meganola
Dyar and the trifines retained in Nola by Poole (1989), and both are
currently becoming assigned to the older name Rhynchopalpus
Hampson by workers such as Inoue (1998, 2001) following Holloway & Miller
(1995). The trifine taxa are reconsidered under Manoba
Walker, and other trifine genera are discussed, particularly
in the Australian and African faunas.

Fig 1.
Tentative phylogeny for the Nolinae as discussed, with indication of possible
apomorphies as follows: A, loss/reduction of first pair of larval prolegs; pair
of scaphial lobes present in male genitalia (also elsewhere in Nolidae); B,
larva with verrucae; C, male antennae bipectinate; D, female with invaginate
signum or signa in bursa; larval head-capsule stacking widespread; E, hindwing
venation trifine; F, partial or complte loss of larval head-capsule stacking.
The classification emerging for the group from current studies, including this
one, is a rather pectinate one as summarised tentatively in Fig 1.
Beana is basal for reasons given earlier, followed by the Barasa
group and Melanographia
Hampson, quadrifines with M3 and CuA1 stalked, primitive corrugate signa and
filiform male antennae. The next sequence of genera is marked by possession of
bipectinate male antennae and development of the invaginated type of signum, the
latter not seen in Dialithoptera, which is therefore placed in the most
basal position, below an unresolved complex of quadrifine genera with modified
signa and male antennae bipectinate. It is amongst these that most records of
larval head capsule stacking occur: the
Acatapaustus Bethune-Baker (= Eurynola
Hampson) complex in the Australasian tropics (Bigger, 1988); Uraba
Walker (=
Coesa
Walker and Toxoloma
Felder) in Australia (McFarland, 1980); Proneca
Swinhoe and Sarbena
Walker (see below); unplaced quadrifine species from the old concept of Roeselia/Meganola
such as gigas
Butler,
mikabo Inoue, nitida
Hampson and
togatulalis H�bner (McFarland, 1980; Sugi, 1987).
Head capsule stacking does not provide a means of defining a natural group of
quadrifine genera or a single genus, as records of it are too sparse, and it
also occurs in Evonima
where hindwing venation is reduced to the trifine condition and male antennae
are filiform, possibly secondarily so, and in one species of
Manoba
where the stacking habit is possibly more vestigial and the capsules less
tightly associated. Thus these quadrifine taxa form an unresolved
polychotomy in Fig 1, and generic placement of such taxa is problematic as there
are no obvious features that permit assignation of quadrifine "Roeselia�
to the genus-group names already available or justify description of new genera,
hence these species are placed in
"Meganola�. Features that may be helpful in resolving this sequence
of quadrifine taxa and its relationship with the trifine sequence discussed next
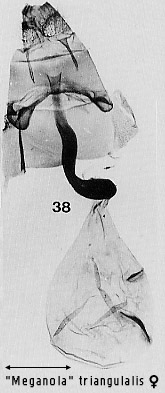
include: a longitudinal arrangement of the two signa in the bursa, seen in
several "Meganola�
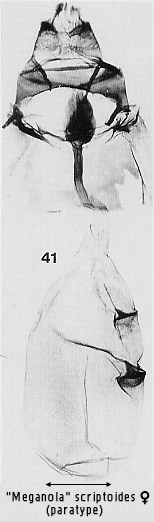
species (Figs 30, 32, 38, 41; see also illustrations for some Meganola
species in Inoue (1982)), but also in Evonima (Fig 37) and brunellus
Hampson and allies in Manoba; a double-peaked area that stains
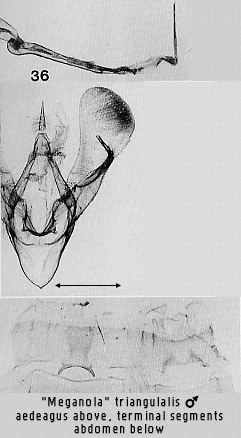
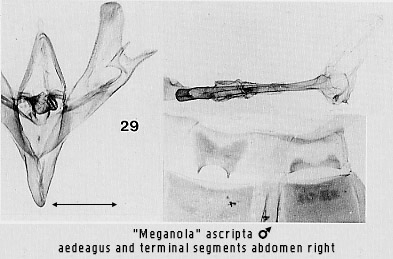
darker with chlorazol black on the basal margin of the male eighth sternite,
seen in Figs 29 and 36.
Evonima,
with head-capsule stacking, is placed as sister-group to the remaining trifine
taxa where weak or no head-capsule stacking has been recorded and where it has
probably been secondarily lost. The relationships of remaining trifine genera
are currently unresolved. Nola can be clearly defined, and a concept of
Manoba
Walker, an older name, is offered below for the bulk of trifine
Rhynchopalpus
/ Meganola
taxa. However, there are additional trifine genera in Australia (Dimona
Walker, Sorocostia
Rosenstock, Tribunta
Walker), Africa (e.g. Metanola
van Son, Paranola
van Son, Poliothripa
Hampson) and the New World (e.g. Callinola
Butler) where the relationships are unclear except they do not fall within the
definitions of Nola
and Manoba
given here.
Limited material of several other Bornean noline species was also examined in
London and from the Zoological Museum, Univ. Amsterdam. These species could not
be assigned with confidence to any of the generic concepts discussed below.
Discussion of them is therefore held over until more material becomes available
and the studies of colleagues in the Hungarian Natural History Museum, Budapest,
and their associates have established clearer generic concepts based on an
appreciation of taxa from throughout the range of the subfamily.
Reference to this research effort and information kindly provided to the author
will be attributed to the 'Budapest group'.
>>Forward
<<Return to Contents page
|