|
The status of the Trisatelini
Beck (1999) proposed the tribe Trisatelini in the Hypeninae on the basis of larval characters, though Owada (1987: 46) had stated that Trisateles Tams was not a herminiine. Sugi (1987) and Randolf & L�dl (2001) noted that the counter-tympanal hood was postspiracular, thus supporting Owada if this feature is accepted as a clear distinction of non-herminiines (but see comments on p. 17). Sugi had earlier (Inoue et al., 1982) tentatively moved the genus to the Acontiinae. Bretherton et al. (1983) retained the genus in a broad concept of the Hypeninae. The tribe was formally synonymised with the Herminiinae by Fibiger & Lafontaine (2005). However, in the same publication, Fibiger & Hacker (2005) listed the genus in the Phytometrinae! Thus, the history of the taxon is confused, and its status is still unclear.
The larva (Baker, 1969; Bretherton et al., 1983; Beck, 1999) has no prolegs on A3 and only vestigial ones on A4. When mature, it has a golden appearance with dark brown mottling and, on the head, reticulation. In the fourth instar the larva appears velvety due to the presence of a fine dense pubescence. Pupation is in a cocoon of fine white silk spun within dead leaves pulled together with prominent struts of silk. The pupal cremaster is reticulated with ridges, further ridges extending anteriorly from this. Within the reticulate area are six small hooks in the basal part and a pair of much larger ones apically. The larvae feed on withered leaves of Quercus but would also take Fagus (Fagaceae).
Sugi (1987), following an earlier suggestion by M. Owada (pers. comm.), drew attention to the similarity between Trisateles and the Japanese Oruza mira Butler in life history, the latter showing all the salient features of the former: proleg reduction, a dense covering of velvety secondary setae; dead leaf feeding. The larva illustrated by Sugi has a similar mottled and striated brown appearance. Similar proleg reduction and pubescence is seen in other species of Oruza Walker and Lophoruza Moore, but there is greater diversity in diet.
Examination of the male and female genitalia of the type species of Trisateles, emortualis Denis & Schifferm�ller, supports the contention of Owada that the genus is not a typical herminiine, and that of Sugi that the genus is related to Oruza mira. The genitalia lack the herminiine features noted above and share features such as: a trifid valve apex of a type not seen in herminiines but present in other Oruza Walker (e.g. kavatcha Holloway; Holloway, 1979: fig 104); a small, rather digitate pocket between each apodeme of the female eighth segment and the ostium bursae, the apodemes themselves being short; uniform sclerotisation, rather than reinforcement by sclerotised bands, of the ductus bursae; small, shallow phragma lobes of the second abdominal tergite. Some of the features noted above are also seen to some extent in the Saroba group of genera discussed by Holloway (2005), and many species of the Oruza complex have the base of the corpus bursae slightly coiled as in that group. A few of these features are also seen in the Aventiinae.
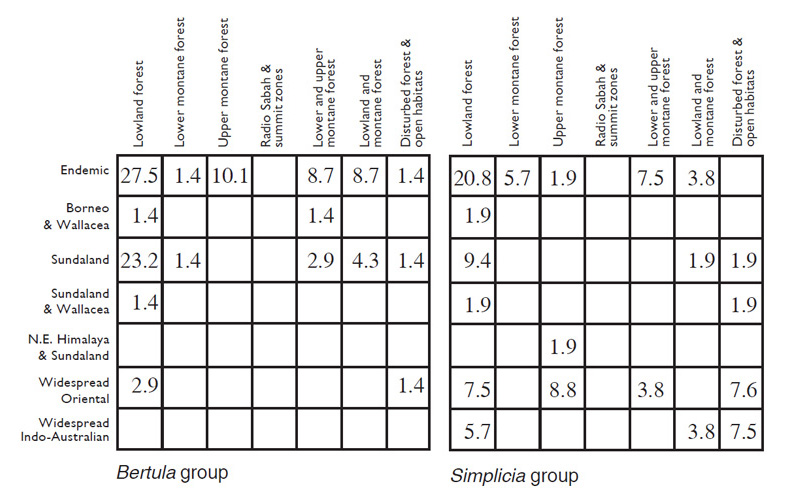
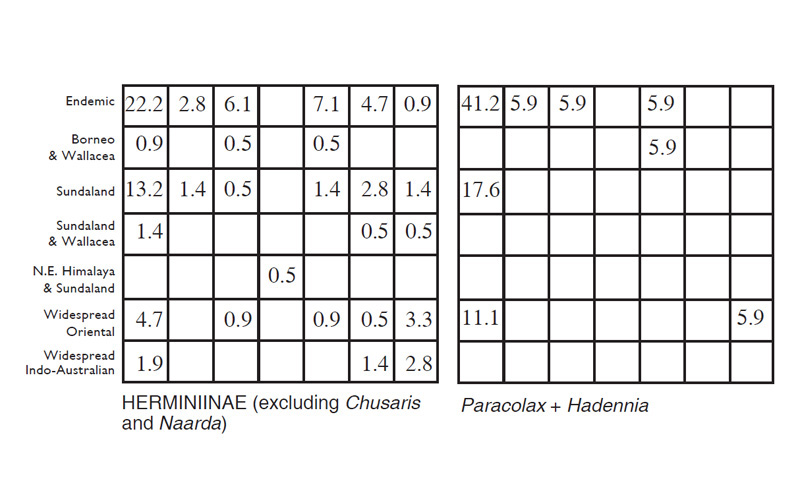
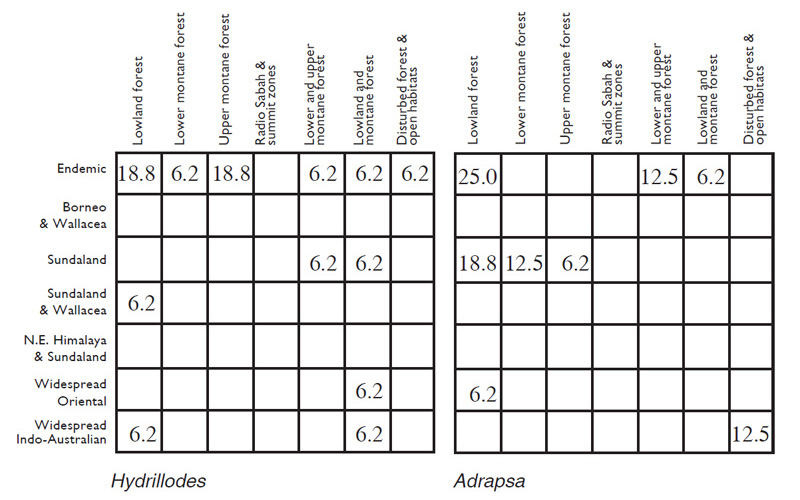
Table 2. Percentage of species for Herminiinae groups of genera and the core subfamily as a whole amongst various biogeographic and ecological categories as discussed in the text.
Thus, the status and relationships of the group of genera around Oruza, currently a traditional acontiine, require investigation and will be considered further in the part of this series treating that complex, including the Aventiinae. The validity of the Trisatelini as a higher taxon will be assessed within this diverse context. At the time of going to press, progress with that part is indicating increasingly clearly that the concept of the Aventiinae should be expanded considerably to include the Oruza complex, genera related to Corgatha Walker, Cerynea Walker and Arasada Moore, the Trisatelini, the Eublemminae (but not the Pangraptini) and the Saroba group.
<<Back
>>Forward
<<Return to Contents page
|