|
This tribe embraces the concept of the Macaridae of Guenée (1857), a
name that should take priority (as Macariini) despite the extensive usage of
Semiothisini, probably first employed by Warren (1894; as Semiothisinae).
Packard (1876) merged Guenée's Macaridae and Zerenidae with his
Fidonidae as the Fidoninae (more correctly Fidoniinae). This is based on the
Fidonites of Duponchel, typified by Fidonia Treitschke. Fidonia is
a junior objective synonym of Eurrhanthis Hübner, having as common type
species the Mediterranean plummistaria Villers. This is distinct from the
concept of Semiothisini of Forbes (1948) as it backs the definitive features
discussed below.
The Macariini also embrace the Fernaldellinae of Hulst (1896), as Fernaldella
Hulst is placed as a semiothisine by McGuffin (1972).
Forbes referred to male genitalia with two setal spines (or horns) at
the tip of the uncus and a deeply emarginate or forked valve. The chaetosemata
are transversely elongate, tapering towards the centre of the head through a
parting of the head scales. The antennae are bipectinate, serrate or filiform
with cilia. The forewing is often foveate in a characteristic manner. There is a transverse comb of setae on sternite 3 of the male.
No single character can be used to define the tribe but a combination of
the two genitalic features mentioned and that of the chaetosemata will enable
members to be identified reliably. The chaetosema character is, however, present
in Scardamia Guenée of the Scardamiini (See
Scardamiini), a probable
homoplasious occurrence given the weight of other characters. The gnathus tends
to be strong, expanded deeply at the apex.
The development of the male forewing fovea requires further study across
the tribe. It is strongly developed in most S.E. Asian taxa that have been
attributed to Semiothisa, and is illustrated in Fig 1. The base of vein
A1 is sigmoid, its anterior convexity containing a corrugate tymbal structure;
this is bounded distally also by a spur from the posterior vein enclosing the
cell (CuA). The vestige of A2 is strongly curved, running parallel to, but
mostly well separate from, the basal curvature on Al. In Macaria and Semiothisa
often only the spur from the cell is evident, but this is variable within
these genera (Hua Baozheng, pers. comm.).
 |
Normal chaetosemata are present in Lampadopteryx Warren, Hypephyra
Butler and Iridoplecta Warren, but the first two have the uncus
horned, and all have the valve forked, albeit weakly in Lampadopteryx. There
is no fovea in these genera, hence its degree of development within the tribe
will in general be apomorphic.
The apomorphic state of the chaetosemata occurs in a number of moderate
to large genera such as Itame Hübner, Semiothisa Hübner, Godonela
Biosduval, Tephrina Guenée and Oxymacaria Warren, as well as
smaller ones such as Chiasmia Hübner, and Parosteodes Warren.
However, the definition and status of these genera is far from satisfactory.
Inoue (1986) indicated that the type species of Semiothisa, the South
American gambaria Hübner, has features that set it apart from the
Holarctic and Old World tropical taxa currently placed in the genus. It has the
costal arm of the male valve slender, curved, apically acute, and lacks uncus
horns. The male antennae are serrate. He placed a number of species lacking
uncus horns in Heterocallia and referred Oriental taxa with horns and
with filiform male antennae to Chiasmia.
In order to resolve this question a little further to determine the
status of Bornean taxa, males of all relevant generic type species (as in
Fletcher (1979)) were dissected. In addition, numerous N. American taxa have
been featured by McGuffin (1972), and there is a review of a N. American conifer
feeding complex (referable to the generic name Sciagraphia Hulst) by
Ferguson (1974).
Important features appear to be the presence or absence of uncus horns,
the state of the distal margin of the eighth sternite, condition of the male
fovea, and, to some extent, whether male antennae are bipectinate or filiform.
Secondary loss of uncus horns and the degree of pectination of the male antennae
are probably somewhat homoplasious, with eighth sternite characters providing
the most useful distinction.
The sternite is unmodified in Heterocallia, Oxymacaria and Parosteodes.
The male antennae are filiform in the first, bipectinate in the second and
serrate in the third. Only in Parosteodes are uncal horns present. In all
three the saccular section of the valve is small, without ornament. Despite the
antennal differences, Heterocallia and Oxymacaria show common
features such as a slightly bifalcate forewing shape, a moderately strong fovea,
and a row of small white marks over the anterior part of the submarginal. There
are thus grounds for bringing them into synonymy . The N. American Trigrammia
Herrich-Schäffer has filiform antennae and genitalic features as in Heterocallia,
but is markedly different in facies characters. Mellilla Grote and Eumacaria
Packard, with bipectinate antennae, also have distinct facies in this
category.
All other genera examined have a pair of processes from the eighth
sternite. These are large, rounded lobes in Semiothisa and in some N.
American taxa referable to Digrammia Gumppenberg (McGuffin, 1972: californiaria
and hebetata groups). In the latter the uncus is apically narrowed,
with the horns vestigial or lost, and there appear to be diagnostic features of
the female genitalia: a bilobed sterigma and loss of the signum typical of most
Macariini. The valves tend to have the saccus portion produced, angular, but the
sternal lobes tend in some taxa towards the status described below. The male
antennae are filiform.
The genera Tephrina, Gnopharmia, Isturgia Hübner and typical Itame (vincularia Hübner) lack uncus horns and have strongly bipectinate male
antennae. The eighth sternite has a pair of processes, usually apically
sclerotised, separated from each other by only a narrow, shallow cleft except in
Isturgia where it is deeper, rounded, and Gnopharmia where
it is broad. The N. American Itame taxa illustrated by McGuffin (1974)
have small uncus horns, a strong valvula (a central lobe between the arms of the
valve) and rather broad sternal processes separated by a strong but narrow
cleft. Their status needs further investigation.
The bulk of taxa currently in Semiothisa should probably be
reassigned to other genera as suggested by Inoue, e.g. Macaria Curtis, as
suggested by Ferguson (in litt.). The extent of the true Semiothisa and
of Macaria in the Neotropics is reviewed by Hua & Scoble (in
preparation).
Macaria (embracing also Philobia Duponchel, Eutropa Hübner and Sciagraphia
Hulst) includes species with sternal processes rather acute, separated by a
deep, basally narrow cleft. The uncus horns are small. There is no valvula, and
the saccular portion of the valve is rounded, with submarginal ornamentation
usually lobes and flanges. The concept embraces many Holarctic (e.g. the ulsterata
and signaria (Sciagraphia) groups of McGuffin) taxa and a number of
Neotropical ones. Male antennae can be serrate or ciliate. The fovea is not well
developed.
Inoue's (1986) concept of Chiasmia includes species with short,
heavily sclerotised sternal processes, separated by a shallow, moderate to broad
cleft (but much reduced in Automolodes and Iulocera Warren). The
male antennae are filiform. The uncus horns are mostly strongly developed. The
saccular process of the valve is diversely developed: it is variably acute,
often ornamented and accompanied by a valvula in the robust species of the Evarzia
Walker (= Gubaria Moore) group, well represented in Borneo. This
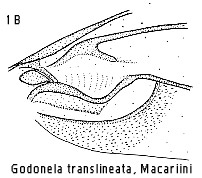
concept embraces the generic names Godonela Boisduval, Acadra Herrich-Schäffer,
Evarzia, Gubaria, Iulocera, Automolodes, Allochrosis Strand and Thyridesia
Wehrli. A facies feature that recurs through the group is an angled forewing
postmedial with a pale mark set just anteriorly and distally to the angle (seen
clearly here in albipuncta Warren). Typical Chiasmia has a ventral
angular process to the costal arm of the valve: a similar feature is seen in the
type species of Allochrosis and in albipuncta).
However, a major problem with this concept of Chiasmia is that
typical Chiasmia lack a fovea on the male forewing, whereas it is very
strongly developed in most Indo-Australian taxa. The facies of typical Chiasmia
is highly modified into a striking reticulate pattern. Hence a more
appropriate generic combination must be sought. Godonela Boisduval is the
oldest name where the type species has a strong fovea, hence this should be
applied to foveate taxa within Inoue's concept of Chiasmia. All the
generic names mentioned above fall within this concept apart from Chiasmia.
Given the paucity of strong apomorphic features, the decisions made here
on placement of species within genera may be subject to revision in future when
a more rigorous, global, morphological survey can be undertaken. Current work on
African macariines by M. Kruger is likely to throw further light on the
situation.
Azata Walker
(type species idriasaria Walker, the type a female without locality data)
was associated by Fletcher (1979) with Indian material related to the Sundanian
species mutabilis Warren and allies , but the facies of the
type specimen is atypical, resembling more S. American taxa in true Semiothisa,
particularly an unnamed series from Jamaica in the NHM (Hua Baozheng, pers.
comm.).
Chiasmiodes Warren Gen. rev. is not a macamine. The male genitalia of the
Himalayan type species variolinea Warren have features more akin to those
of Lithinini.
Eggs and larvae of N. American taxa are described by McGuffin (1974).
Arboreal feeding is general. Pupation is at the surface of the soil in a slight
cocoon. Host-plants are diverse in temperate regions, including several
conifer-feeding taxa or groups. In the tropics there is some preference for
Leguminosae, particularly Acacia, Albizia and allies.
>>Forward
<<Return to Contents page
|