|
The Ennominae are the largest subfamily of the Geometridae. In the
Bornean fauna just under half the species are ennomines. The traditionally
diagnostic feature for the subfamily is loss (or reduction to a fold) of vein M2
on the hindwing. No other features have been located that are shared by all
major tribal groupings within the Ennominae, so the monophyly of the group is
only weakly supported.
The group as a whole has a wide ecological range, occurring in diversity
at all except very high latitudes and, in the tropics, altitudes. In Borneo,
Sulawesi and Seram, ennomines show their greatest diversity in the lower montane
zone (Holloway, 1984a, 1993; Holloway, Robinson & Tuck, 1990).
Some ennomines show specificity to lowland forest types, such as heath
forest, alluvial forest or mangrove. The subfamily also includes several species
that appear to fly predominantly in the forest understorey. Ennominae are
therefore highly suitable as an environmental indicator group except for poor
representation in open habitats (Holloway, 1984a, 1985; Holloway &
Barlow, 1992).
An attempt is made here to define tribal groupings and suggest a higher
classification for the subfamily, though this is based primarily on groups
represented in S.E. Asia. Host-plant specialism is also reviewed in relation to
this classification.
Family-group names
The attempt to assess the higher classification of the Ennominae and to
assign genera to tribal groupings necessitated an investigation of prior usage
of family- group names. This in turn led to the accumulation of about 150
family-group names used for the Geometridae and some indication of when these
names first appeared. For the subfamilies Oenochrominae, Geometrinae, and
Sterrhinae the listings of Prout (1912, 1913, 1934-5) provide a foundation,
though a number of family-group names have been erected subsequently, e.g. in
the Geometrinae by Inoue (1961) and Ferguson (1969). Prout did not publish lists
for the Larentiinae and Ennominae.
In the Code of Zoological Nomenclature, rules of priority for
family-group names tend to apply, but somewhat less strictly than for
genus-group and species-group names. After 1960, family-group names based on
junior generic synonyms may not be replaced by a new name based on the senior
generic synonym, but can be replaced by extant family group names based on
senior generic synonyms or by senior family group names based on other genera
within the concept of the reclassification. Before 1961, such replacements can
stand if the replacement name has won general acceptance. Thus Bistonini should
probably stand in relation to the older Amphidasini and Hyberniini (replaced by
Erannini), though Seven (1991) uses Amphidasini (see list below for details).
Reinstatement of the genus Macaria Curtis from synonymy with Semiothisa
H�bner strengthens the case for preferring Macariini to Semiothisini (as
does Seven), as Macariini is the senior name.
It became apparent during this investigation that reclassification of
the Sterrhinae, a subfamily name indicated by Fletcher (1979) to be preferred,
will probably lead to even greater changes to family-group names. Indeed,
Mikkola et al. (1985) and Seven (1991) have reverted to using
Scopulinae for this subfamily based on the oldest name (Scopulites, Duponchel
(1845)). The situation is therefore reviewed briefly here to exemplify
the problems that can be encountered. Revision of the classification may show
that the group is polyphyletic rather than monophyletic: even if monophyletic,
it may contain seven groupings:
Scopulini (= Acidaliini Duponchel (1845) based on a junior
homonym of a genus-group name outside the Lepidoptera). Scopula and
allied genera;
Cyclophorini Moore (1887) (= Zonosomini White (1876, Scot. Nat. 3:
361), reintroduced probably on priority grounds by Seven (1991) but Cyclophora
is a senior objective synonym of Zonosoma; Cosymbiini Prout (1911, Tr.
City Lond. ent. soc. 20: 23)), which traces back to the Cyclophorae
of H�bner. The name Ephyrini of Guen�e (1857) is based on a junior homonym of
a genus group name outside Lepidoptera but was applied to the concept of
Cyclophorini. Another reason for preference of Zonosomini is the usage of the
family-group name Cyclophoridae in Mollusca, based on Cyclophorus Montfort.
The moth group includes genera with a distinctive girdle to the pupa (Holloway,
Bradley & Carter, 1987);
Idaeini Butler (1881) (= Sterrhini Meyrick (1892), a seniority that must
stand both on grounds of priority and because Sterrha H�bner is
currently treated as a junior subjective synonym of Idaea Treitschke);
Timandridi Stephens (1850) (= Calothysanini Herbulot (1962-3)). This
name is used by Hodges et al. (1983) and Seven (1991);
Rhodometrini Agenjo (1952);
Cyllopodini Warren (1895);
Rhodostrophini Prout (1934-5).
The family-group names (over 60) listed below have been located for
possible usage within the Ennominae. Invalid names, based on junior homonyms,
usually of genus-group names outside the Lepidoptera, are indicated by square
brackets.
The literature references given are only of those considered to indicate
both early usage and currency in modern literature. Whilst an attempt has been
made to ensure the earliest reference has been located, this cannot be taken as
categorical as the search was not exhaustive. The task is considerably
complicated by both past and present reluctance of authors to indicate authority
of the family-group names they use or to own up to new concepts.
However, the earliest dates located for names within Oenochrominae,
Geometrinae and Sterrhinae up to the Catalogus Lepidopterorum publications
of Prout (1912, 1913, 1934-5) were found to be the same as in Prout�s
lists. Many of Duponchel's names are attributed to Guen�e, but represent the
first published occurrence.
ABRAXINI: Abraxinae Warren (1893); Swinhoe (1900), Hodges et al. (1983),
Seven (1991). (See Abraxini.) Conceptually replaced the unavailable
Zerenini, and see
also Pantherini (q.v.).
AMPHIDASINI: Amphidasites Duponchel (1845); Bruand (1846), Guen�e
(1857). Amphidasis Treitschke is a junior objective synonym of Biston Leach
(Fletcher, 1979). Used for concept of Bistonini by Seven (1991). (See
Boarmiini.)
ANAGOGINI: Anagogini Forbes (1948); Hodges et al. (1983).
Subordinated here to Hypochrosini.
ANGERONINI: Angeronini Forbes (1948); Hodges et al. (1983).
Applicable to concept of Aspilatini (q.v.) as Aspitates and Angerona share
a costal ampullate process to the valve and other features of the male genitalia
(see McGuffin (1987)) and Higher Classification.
ASCOTINI: Ascotinae Warren (1893). Included in the strict concept of
Boarmiini of Sato (1984a).
[ASPILATINI]: Aspilatites Duponchel (1845); Bruand (1846). Based on a
misspelling of the genus-group name Aspitates Treitschke (see Fletcher
(1979)). The concept is currently included in Angeronini (q.v.).
AZELININI: Azelinini Forbes (1948); Hodges et al. (1983). Larval characters suggest a relationship to Nacophorini
(Heitzmann, 1985).
BAPTINI: Baptini Forbes (1948); Hodges et al. (1983) include the
older name Palyadini (q.v.) within their concept, but see discussion of Baptini.
BISTONINI:
Bistonidi Stephens (1850); Humphreys (1858-9), Hodges et al. (1983) but
see Amphidasini and Boarmiini, Biston
Leach.
BOARMIINI:
Boarmites Duponchel (1845); Bruand (1846), Stephens (1991). (See
Boarmiini.)
BRACCINI:
Braccinae Warren (1894). (See Boarmiini.)
BUPALINI:
Bupalini Herbulot (1962-3); Leraut (1980). Not investigated here, but the bifid
cremaster indicates relationship to the Boarmiini group of tribes.
CABERINI:
Caberites Duponchel (1845); Bruand (1846), Stephens (1850), Guen�e (1857), Hodges
et al. (1983), Seven (1991). See also Deiliniini (q.v.), and concept may
also incorporate Erastriini and Catopyrrhini (q.v.). (See
Tribe Caberini).
CAMPAEINI:
Campaeini Forbes (1948); Hodges et al. (1983), Seven (1991). Metrocampini
(q.v.) is an older name but based on a junior objective synonym of Campaea Lamarck
(Fletcher, 1979).
CASSYMINI:
This volume. May embrace the N. American (Forbes, 1948) concept of Abraxini (See
Tribe Cassymini).
CATOPYRRHINI:
Catopyrrhinae Warren (1894). See Erastriini (q.v.) and Tribe
Caberini.
CHEIMATOBIINI:
Cheimatobiidi Tutt (1896). See Theriini (q.v.). Cheimatobia Stephens is a
junior objective synonym of Theria H�bner.
CINGILIINI:
Cingiliini Forbes (1948). Some usage in N. America but generally subordinated to
Ourapterygini (q.v., Forbes et al. (1983)). The relationship between the
two needs clarification; larval characters indicate a close
relationship with the Ennomini (Heitzmann, 1985).
CLEORINI:
Cleorites Duponchel (1845); Stephens (1850). Included in the strict
concept of Boarmiini of Sato (1984a).
COLOTOINI:
Colotoini Herbulot (1962-3); Seven (1991).
COMPSOPTERINI:
Compsopterini Herbulot (1962-3); Leraut (1980). See Ligiini, Pachycnemiini and
Prosopolophiini (q.v.). Status not investigated here. The type genus includes a
number of robust Palaearctic species.
CROCALLINI:
Crocallidi Tutt (1896). No current usage located.
DASYDIINI:
Dasydites Duponchel (1845); Stephens (1850), Tutt (1896). Based on a junior
objective synonym of Sciadia H�bner. Concept not investigated here.
DEILINIINI:
Deiliniinae Warren (1893). (See Tribe Caberini).
EMPLOCIINI:
Emplocidae Guen�e (1857). Based on a Neotropical genus; not investigated here.
ENNOMINAE:
Ennomites Duponchel (1845); Bruand (1846), Guen�e (1857), Hodges et al. (1982),
Seven (1991). See also Odopterini (q.v.) and Cingiliini.
EPIONINI: Epionidae Bruand (1846); Stephens (1850), Humphreys (1858-9).
EPIRRHANTHINI: Epirrhanthini Forbes (1948); Hodges et al. (1983).
Not investigated. McGuffin (1981) notes some similarities with Lithinini (q.v.).
A small N. American group.
ERANNINI: Eranniinae Tutt (1896); Dugdale (1961). See also Hyberniini
(q.v.). Hybernia Berthold is a junior objective synonym of Erannis H�bner
(Fletcher, 1979). Tutt included Eranniinae as a subfamily of Hyberniidae! Taxa
usually included in modern concepts of Bistonini/Amphidasini.
ERASTRIINI: Erastriae H�bner (1825); Erastridae Herrich-Sch�ffer
(1845); Stephens (1850). The original concept was applied to the acontiine
Noctuidae for a considerable time. Classification of the generic type species
(see Fletcher, 1979) leaves this family-group name applicable within the
Ennominae. See Cryptopyrrhini and Tribe Caberini.
EUBYJINI: Eubyjinae Warren (1893). Eubyja H�bner is based on a
species currently included in Biston Leach. See Amphidasini and Bistonini.
EUTOEINI: This volume; (See Eutoeini).
FERNALDELLINI: Fernaldellinae Hulst (1896) Based on a N. American genus
currently placed within the concept (Hodges et al. 1983) of the Macariini/ Semiothisini (q.v. and
see Macariini).
FIDONIINI: Fidonites Duponchel (1845); Bruand (1846), Stephens (1850),
Guen�e (1857). Has been used to include Macariini/Semiothisini
subsequently (Packard, 1876) but is based on an unrelated S. European taxon. (See
Macariini). The name has not been used by Herbulot (1962-3) or Leraut (1980).
GNOPHINI: Gnophites Duponchel (1845); Bruand (1846), Stephens
(1850), Seven (1991). Not investigated in detail but probably subsumed by the
concept of Boarmiini in this volume.
GONODONTINI: Gonodontini Forbes (1948); Hodges et al. (1983).
Applied in the sense of Odontoperini (q.v.). Redefined in this volume (See
Gonodontini).
HYBERNIINI: Hibernites Duponchel (1845); Bruand (1846), Stephens (1850),
Guen�e (1857). See Erannini. Falls within current concepts of Amphidasini/
Bistonini.
HYPOCHROSINI: Hypochrosidae Guen�e (1857); Kirby (1897). See
Anagogini and (See Hypochrosini).
LACARIINI: Lacarini Orfila & Schajovskoy (1959). Associated with
Lithinini by these authors and Rindge (1986). (See
Lithinini).
[LIGIINI]: Ligidae Guen�e (1857). Based on a junior homonym of a genus group name outside
Lepidoptera. See Compsopterini, Pachycnemiini and
Prosopolophini (q.v.).
LITHININI: Lithinini Forbes (1948); Hodges et al. (1983). (See
Lithinini).
MACARIINI:
Macaridae Guen�e (1857); Moore (1887), Swinhoe (1900), Seven (1991). See
Semiothisini (q.v.), Fernaldellini and Macariini.
MELANCHROIINI:
Melanchroiinae Hulst (1896); Kirby (1897). Applicable to a small New World
group; not investigated here.
MELANOLOPHINI:
Melanolophini Forbes (1948); Hodges et al. (1983). A New World group that
probably falls within the broad concept of Boarmiini applied here.
METROCAMPINI:
Metrocampidae Tutt (1896). See Campaeini.
MILIONIINI:
Milioniini Inoue (1992) is the only reference so far located. In the absence of
a published definition, or mention before 1930, the name is not available. The
type genus falls within the concept of Boarmiini here (See
Boarmiini).
NACOPHORINI:
Nacophorini Forbes (1948); Hodges et al. (1983). A New World group
outlined by Rindge (1961). The tribe is possibly related to the Azelinini (see
above).
NEPHODIINI:
Nephodiinae Warren (1894). A Neotropical group not studied here.
OBEIDIINI:
Obeidiini Inoue (1992) is the only reference so far located. In the absence of a
published definition, or mention before 1930, the name is not available. The
type genus falls within the concept of Boarmiini here (See
Boarmiini
).
ODONTOPERINI:
Odontoperinae Tutt (1896). Fulfils the N. American concept of Gonodontini (see
above and (See Gonodontini).
[ODOPTERINI]:
Odopteridi Stephens (1850), Humphreys (1858-9). Odoptera Sodoffsky is an
unnecessary objective replacement name of Ennomos Treitschke. See
Ennominae.
ONYCHORINI:
Onychorini Herbulot (1962-3); Leraut (1980). Based on W. Palaearctic genus not
studied here.
OURAPTERYGINI:
Urapteridae Bruand (1846); Guen�e (1857), Warren (1893), Hampson (1918;
preferred to Ennominae for subfamily, as based on the oldest genus-group name),
Forbes (1948; ?first spelling as Our- rather than Ur-), Hodges, et al. (1983).
See Cingiliini and Ourapterygini.
PACHYCNEMIINI:
Pachycnemiidae Kirby (1903); as replacement name for Ligidae. See above and also
Compsopterini and Prosopolophini.
PALYADINI:
Palyadae Guen�e (1857); Moore (1887), Warren (1894), Hulst (1896), Kirby
(1897). Subordinated to Baptini in Hodges et al. (1983) but see Baptini.
Guen�e's concept of this group is valid today if Eumelea Duncan is
excluded (M.J. Scoble, pers. comm.).
[PANTHERINI]:
Pantheridae Moore (1887), as replacement for Zerenini (q.v.) but Panthera H�bner
is a junior homonym of a genus-group name outside the Lepidoptera (Fletcher,
1979). (See Abraxini.)
PLUTODINI:
Plutodinae Warren (1894); Swinhoe (1900). (See
Plutodini).
PROSOPOLOPHINI:
Prosopolophinae Warren (1894); Swinhoe (1900). Proposed to replace Ligidae (see
above, and also Compsopterini and Pachycnemiini).
RUMIINI: Rumiinae
Tutt (1896). Based on Rumia Duponchel, a junior objective synonym of Opisthograptis
H�bner. No current usage located.
SCARDAMIINI:
Scardamiinae Warren (1894). (See
Scardamiini).
SELENIINI: Seleniidi
Tutt (1896). Probably best subsumed by Ennomini; (See
Introduction).
SELIDOSEMINI:
Selidosemidae Meyrick (1892); Warren (1893). Based on a European and N. African
genus not studied here. The concept of earlier authors probably coincides with
Boarmiini as treated here.
SEMIOTHISINI:
Semiothisinae Warren (1894); Hodges et al. (1983). See Macariini and
Fernaldellini.
SIONINI: Sionites
Duponchel (1845); Bruand (1946), Stephens (1850), Guen�e (1857). Based
on a European species included in Gnophini by Herbulot (1962-3).
SPHACELODINI: Sphacelodini Forbes (1948); Hodges et al. (1983). A New World group not
investigated here.
THERIINI: Theriini
Herbulot (1962-3); Leraut (1980). See Cheimatobiini. This is based on a
Palaearctic genus that, from pupal cremastral characters, probably falls within
the broad concept of Boarmiini applied here.
THINOPTERYGINI:
This volume. (See Thinopterygini )
[ZERENINI]:
Zerenites Duponchel (1845); Bruand (1846), Stephens (1850), Guen�e (1857). Based
on a junior homonym of a genus-group name outside the Lepidoptera. See Abraxini
and Pantherini.
The higher
classification of the Ennominae
An attempt has been
made in the systematic section to group Bornean genera of ennomines into tribes.
In some instances (e.g. Caberini, Baptini) it was possible to recognise a major
generic grouping defined by several characters, but association of this grouping
with the type genus of the tribe was more tenuous.
The absence as a
tubular vein or weakening of vein M2 in the hindwing currently defines the
subfamily. In this section potential groupings of tribes within it will be
assessed.
Forbes (1948)
divided N. American tribes on characters of the pupal cremaster: whether it was
apically bifid only (Group D of Mosher (1916)) or with more than two spines
(usually eight: four pairs). McGuffin (1981) suggested that the bifid condition
was the more primitive, reversing an earlier opinion which, on surveying
cremaster characters in Geometroidea generally, was probably correct: the bifid
condition is derived.
Data on geometroid pupae from Forbes, McGuffin, Mosher (1916), Sato
(1984a), Carter (1984), Khotko (1977) and the manuscript notes on the Indian
fauna by T.R.D. Bell indicate that possession of eight cremastral hooked shafts
is the commonest and more generally distributed condition seen in the
Cyclophorini, Naxa Walker amongst the Oenochrominae (?Orthostixini), the
majority of Geometrinae and Larentiinae, and also in the Drepanidae, Uraniidae
and Epiplemidae.
The bifid condition is not unique to the Ennominae but is much less
frequent elsewhere. It occurs in some, but not all, Scopulini where the
cremaster consists of two parallel shafts rather than divergent ones. The
Alsophilini have the cremaster bifid, almost T-shaped as does Operophtera (but
not Epirrita, where the 'T' is associated with two pairs of reduced
hooklets) of the larentiine tribe Operophterini (Packard, 1876; = Oporiniini of
Seven (1991)) (Carter, 1984). In the Eupitheciini all conditions occur from
eight shaftlets, through two strong ones and six weaker ones, to two shaftlets;
the Epiplemidae show a similar range of conditions. The bifid condition is seen
in some other larentiine genera such as Aplocera Stephens and Lithostege
H�bner. In the Archiearinae the cremaster resembles that of Alsophila (Mosher,
1916; Nakamura, 1987).
Hence it is probable that, within the Ennominae, the divergently bifid
cremaster represents a derived condition. It brings together the Boarmiini sensu
lato (See Boarmiini) with the
Macariini, Cassymini and Eutoeini. The inclusion
of Abraxini by Forbes rests on his examination of N. American genera he assigned
that tribe but which are probably better placed in Cassymini (See
Cassymini n.).
Adults
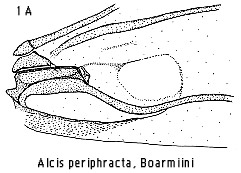
All the tribes with a bifid cremaster have the male forewing foveate in
a high proportion of genera, although the fovea is structurally different in
each (Fig 1). Occurrence of a forewing fovea is rare elsewhere in the Ennominae,
e.g. in Corymica Walker of the Hypochrosini and Borbacha Moore,
tentatively assigned here to the Baptini. Debauche (1937) defined a subgeneric
taxon in the Palyadini on presence of a fovea, somewhat as in Borbacha, but
with a spur from vein CuA as in the Macariini. Outside the Ennominae a fovea
occurs in Noreia venusta Warren, uniquely in a pantropical complex of
genera currently in the Oenochrominae that was identified by Holloway (1984b:
139). This complex must now be expanded to include the Indo-Australian genus Celerena
Walker as it shares the definitive features of the male abdomen. The
geometrine genus Dysphania H�bner also has a strong fovea, but again
structurally different from ennomine types.
Nevertheless, the frequency of the foveate condition supports the
relationship of these ennomine tribes suggested by the pupal character. A
further character that may serve to unite them is the presence of a cucullate
anterior apical portion of the valve that supports a concentration of the valve
setae; these setae extend more sparsely down the costal zone of the valve. In
the Macariini and Eutoeini this zone is separated from the sacculus as the
dorsal arm of the valve by a c!eavage that extends almost to the base of the
valve. In the Cassymini the valve is similarly cleft but the dorsal arm is
reduced to a slender process that usually only bears a few apical setae. In
other tribes the setae tend to be more generally and sparsely distributed over
the anterior of the valve lamina or, as in the Palyadini and a generic grouping
in the Baptini, concentrated elsewhere on the valve. However, not
all Boarmiini exhibit this cucullus.
The division of the valve may provide a synapomorphy to group the Cassymini, Eutoeini and
Macariini, but it is not clear whether the mode of
cleavage is homologous or homoplasious: it appears somewhat different in each
case.
Another feature common to all four tribes is the presence in many genera
of a transverse comb of posteriorly directed setae on the male third abdominal
segment (e.g. Figs 232, 251, 296-8, 349, 369, 494). This appears often to be
correlated with the presence of a hair pencil sheathed within the hind tibia
(e.g. Rindge, 1983) and may serve as a means of distributing scent from that
pencil. But this feature is also seen in other tribes, e.g. for the Bornean
fauna: Abraxini, Gonodontini, Lithinini, Ourapterygini.
Setal ornamentation is seen elsewhere in the Geometridae but not in the
form of a transverse comb. In the Noreia/Celerena group of the
Oenochrominae mentioned earlier the setae occur in two robust patches arranged
symmetrically on either side of the mid-line, separated by a group of finer
setae forming together a roughly circular area (Celerena) or in two
sparser, more widely separated patches with no setae in between (Noreia Walker).
In the Geometrinae there is usually a diffuse patch on each side linked by a
region with sparser setae in between. These are more easily detached than in
Ennominae, except in Pingasa Moore and allied genera where they are more
robust.
In the Noreia group and Ozola Walker (Desmobathrini) in
the Oenochrominae and in the Geometrinae, occurrence of a tibial hair pencil is
frequent but the transverse comb arrangement of the sternal setae appears to be
unique to the Ennominae. The occurrence of a hair pencil/setal connection in
groups outside the subfamily makes it unclear whether the comb is a feature of
the Ennominae ground plan, and therefore its consistent absence in a tribe is a
derived feature, or whether it can be treated as a synapomorphy for the tribes
that possess it, i.e. grouping the Abraxini, Gonodontini, Lithinini and
Ourapterygini with the bifid cremaster complex of tribes.
Rindge (1983) treated presence of setal comb and hair pencil as
apomorphic states within his concept of the Nacophorini, a New World tribe with
some representation in Australia (McQuillan, 1981) that has variable pupal
cremastral ornamentation from two to eight shaftlets, and was assigned by Forbes
(1948) to his non-bifid group of tribes.
McQuillan (1985) revised the Australian ennomine genus Mnesampela Guest,
where the larvae have a full set of prolegs albeit with those on A3 somewhat
reduced; it was suggested by McQuillan to be one of a number of rather
generalised Australian taxa. The pupa has four cremastral hooks and the male
genitalia have socii and a furca (see below). There is a setal comb on the male
third sternite and a hind-tibial hair pencil. This may indicate that the
hypothesis of presence a setal comb being a relatively basal feature in the
ennomine phylogeny and subsequent loss being the derived state is correct.
Consistent absence of a setal comb throughout a tribe, perhaps
associated with loss of a tibial hair pencil, might serve to unite a number of
robust-bodied tribes around the Ennomini, including here the Hypochrosini and
Scardamiini. These share with the Epionini extension and looping of the vinculum
to support lateral coremata. Other tribes recognised in the Palaearctic and N.
American faunas may associate with such a group, such as the Angeronini,
Colotoini, Crocallini and Seleniini.
Many of these groups have a strong furca arising between the bases of
the valves in the male genitalia, but a similar feature is seen in Ourapterygini
where a setal comb is present, and some Boarmiini. Anellar processes, or
processes from the juxta, probably not homologous with the furca in these
tribes, occur in the Lithinini and Nacophorini.
Presence of socii associated with the uncus is probably a primitive
feature as they also occur in Geometrinae. In the Ennominae they are much
reduced or absent in the 'bifid cremaster' group of tribes and also the Abraxini, Thinopterygini and
Gonodontini.
In the female genitalia a single, often mushroom-shaped, usually spined
or dentate signum is more or less general to the subfamily (Fig 2). It is
modified into a ridged structure in Lithinini. More general scobination with no
signum is seen in the Plutodini and the Campaeini (or Metrocampini).




Larvae
Study of the larvae of Ennominae has probably not yet been comprehensive
enough to provide any clear pointers to higher classification. Sato (1984a)
suggested that presence of four external setae on the A6 proleg was apomorphic
for his Hypomecis group, but this characteristic was regarded as
diagnostic for the subfamily relative to Geometrinae by Singh (1953), though not
exclusive to it, and is a feature of the putative primitive ennomine genus Mnesampela
(McQuillan,1985). In Geometrinae the proleg has three external setae.
Dugdale (1961) compared New Zealand ennomine larvae with published
accounts from the Nearctic (earlier papers by McGuffin) and India (Singh, 1953).
He identified four characteristics common to ennomines treated in these works
and possibly diagnostic: (1) anal shield with seta SD1 anterior to seta D1; (2)
hypoproct produced to a fine point, strongly developed; (3) four to eight
annulets (transverse sclerotised bands that are muscle attachments) on A1 -5;
(4) SV group tri- (to 6) setose on A2-5 (cf. bisetose).
Heitzman (1985) performed cladistic analyses on 70 larval characters for
genera of Nearctic Ennomini and related tribes. He found no apomorphies for the
Ennominae as a whole but defined the Ennomini (including the Cingiliini and the
N. American concept of Ourapterygini) on possession of the V2 primary seta. He
noted more than four external SV setae on the A6 proleg in Angeronini,
Epirranthini and Seleniini as well as in the groups referred to below.
Heitzman (1985) and Poole (1987) indicated that the Azelinini (Pero
Herrich-Sch�ffer) and Nacophorini shared a unique arrangement of multiple
setae on the proleg on A6 (he illustrated 16 setae). Singh recorded up to 16
setae on the proleg of Odontopera similaria Moore (as Gonodontis) so
the Odontoperini may be related to the other two tribes. The Lithinini appear to
be characterised by the presence of six external setae (Heitzman, 1985; J.
Weintraub, pers. comm.)
The Scardamiini and Hypochrosini studied by Singh had five external
setae on the proleg, support for the relationship of these groups suggested
above from consideration of the vinculum modification. Five external setae are
found also in the larva of Traminda Warren in the Sterrhinae (Singh). The
group of genera including Anagoga indicated by Heitzman to be closely
related are referable to Hypochrosini. His other anagogine genera may be
misplaced.
Eggs
Salkeld (1983) surveyed the eggs of Canadian Geometridae. Amongst the 'bifid cremaster' grouping, particularly the Boarmiini sensu lato, there
is a tendency for the walls of the polygonal cells ornamenting the surface to be
heavy and for the cells to be aligned in longitudinal rows. In the putative
Cassymini ('Abraxini') the wavy polygonal cell walls are overlain by a
prominent reticulum, reminiscent of some Larentiinae. The Macariini appear to be
a homogeneous group, with eggs of a rugged appearance, the polygonal cells with
broad and elevated walls, sometimes enhanced by large, domed aeropyles at the
wall junction and a roughly pitted texture to the chorion.
A columnar arrangement of polygonal cells, leading to the walls
separating adjacent rows forming longitudinal ridges is seen in Lithinini and
Caberini. In the former the eggs are not attached to the substrate.
A thick rolled collar encircling the anterior pole is recorded from
several genera in the Ennomini, Hypochrosini (as Anagogini) and Ourapterygini
(including Cingiliini).
Conclusions
The features of adults and early stages just discussed suggest
phylogenetic relationships amongst ennomine tribes somewhat different to those
suggested by McGuffin (1987). His basal groups can be brought together if the
bifid cremaster is considered derived, supported by fovea and valve cucullus
features, as well as loss of socii. This would include a broad concept of
Boarmiini (embracing Bistonini and probably Melanolophiini) together with
Macariini, Cassymini (his Abraxini) and Eutoeini (an Old World group defined
here). True Abraxini and Gonodontini may be related to that group (absence of
socii), and Thinopterygini can also be tentatively placed here, though the setal
comb has been lost.
Loss of setal comb, presence of a strong furca and occurrence of
collared eggs could bring together Hypochrosini (Anagogini), Scardamiini,
Epionini and perhaps Ennomini, Angeronini, other small Holarctic tribes, with
Ourapterygini (+ Cingiliini) also associated though a setal comb is present. The
first two tribes group on larval characters, and the others also share
possession of additional SV setae on the A6 proleg, a feature developed to an
extreme in the next group.
The Nacophorini, Azelinini and Odontoperini appear to form a group on
the basis of larval characters. None is represented in Borneo.
The Lithinini and Caberini may group on features of the egg. The Baptini
and Plutodini appear relatively isolated, though a grouping in the former shows
parallels with the Neotropical Palyadini.
The tribes are therefore treated in the systematic section in the order:
Hypochrosini, Scardamiini, Ourapterygini, Baptini, Plutodini, Lithinini,
Caberini, Thinopterygini, Gonodontini, Abraxini, Cassymini, Eutoeini, Macariini,
Boarmiini (sensu lato). The assignation of genera to these tribes is
based as far as possible on an appreciation of shared, derived features
(synapomorphies): it therefore differs in many respects from the more
traditional arrangement of Inoue (1992).
Host-plant specialisation
The majority of ennomines are arboreal feeders as larvae. Host-plant
specialisation ranges from extreme polyphagy in some tribes to restriction to a
particular plant family in others.
The Hypochrosini are broadly polyphagous but with some specialisation at a
generic level, e.g. Heterolocha Lederer on Caprifoliaceae and the
sister-genera Fascellina Walker and Corymica Walker on Lauraceae
and Illiciaceae. The few records for Achrosis Guen�e are from the
Rubiaceae.
The Scardamiini show some association with Flacourtiaceae.
The Ourapterygini are polyphagous.
In the Baptini, the typical genus Lomographa H�bner is a Rosaceae
feeder, but the complex of genera making up the majority of the Bornean fauna in
the tribe is associated with the Aquifoliaceae and, in one instance, the
Flacourtiaceae. There are some records from Piper (Piperaceae).
No records for the Plutodini have been located.
The Lithinini are fern specialists. Indeed, Entomopteryx Guen�e was
associated tentatively with the tribe on the basis of adult morphology in an
early draft of the text, the placement has been supported by biological
information discovered between then and going to press (See
Lithinini et seq.).
The Caberini are strongly associated with the Rhamnaceae, apart from the
type genus which feeds on Betulaceae and Salicaceae.
The only records for the Thinopterygini are from Vitaceae.
The Gonodontini and Abraxini are polyphagous.
Some Cassymini genera show preference for Leguminosae, but there is one record
from Guttiferae. Possible Palaearctic representatives are more polyphagous.
The few records for the Eutoeini suggest a preference for Melastomataceae.
The Macariini as a whole are polyphagous, with some temperate groups specialist
on conifers, but the tropical representatives show a strong preference for
Leguminosae.
The Boarmiini include some highly polyphagous genera but also a number that are
much more narrowly specialised. Milionia Walker is only recorded from
conifers, particularly southern hemisphere genera such as Araucaria,
Dacrydium and Podocarpus. Several genera show some preference for the
Lauraceae, such as Racotis Moore, Xandrames Moore, Amblychia Guen�e
and the related pair, Krananda Moore and Zanclopera Warren. Racotis
has been recorded from the Annonaceae and Amblychia from the
Illiciaceae (also Pogonopygia Warren), families distantly related to the
Lauraceae within the Magnoliidae. Two genera, Amraica Moore and Microcalicha
Sato, are associated with the Celastraceae.
Ecology and geography
The Ennominae are probably the most diverse and generally distributed of the
geometrid subfamilies. The Geometrinae have a strong tropical focus, whereas the
Larentiinae are more diverse in temperate latitudes and at high altitudes in the
tropics (Holloway, 1986, 1993). In Borneo the greatest diversity of ennomines is
in the lower montane zone at around 1000m where an association of species
specific to that zone overlaps with the upper reaches of the lowland forest
fauna and the lower reaches of an upper montane zone fauna. The Ennominae are
particularly strongly represented within this lower montane association.
The subfamily also has a higher incidence of specificity to particular types of
lowland forest, such as kerangas (heath) forest types or that on limestone.
Examples of the former include Achrosis longifurca sp. n., Achrosis
kerangatis sp. n., Synegia botydaria Guen�e, Synegia eumeleata Walker,
Bulonga schistacearia Walker, Peratophyga sobrina Prout (also
montane forests), Sundagrapha lepidata Prout and Diplurodes kerangatis
sp. n.
Limestone specialism is exemplified by Achrosis calcicola sp. n., and Achrosis
alienata Walker.
There are two definite mangrove specialists: Gonodontis clelia Cramer and
Cleora injectaria Walker.
Whilst there are no purely open habitat species, there are a number that
respond positively to forest disturbance and are probably associated with plant
taxa characteristic of regenerating forest. Hypochrosis binexata Walker, Hypochrosis
pyrrhophaeata Walker, Bracca georgiata Walker and Craspedosis
arycandata Guen�e were identified as such by Holloway, Kirk-Spriggs &
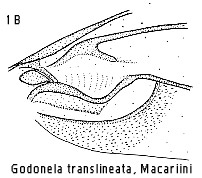
Chey (1992), and some Cleora Curtis, Hyposidra Guen�e and Godonela
Boisduval species may fall into this category too.
Just over 40% of the species are characteristic of the lowlands, and just under
40% are montane, with the remainder being recorded over a wide altitude range.
About a quarter of the lowland species are endemic whereas this proportion
increases to a half for montane species, the overall proportion of endemics
being 30%. A third of all ennomines consists of species restricted to land on
the Sunda Shelf with a further 6% extending from Sundaland to the Philippines or
Sulawesi. About a quarter are more widely distributed in the Oriental tropics.
Only about 5% range more widely through the Indo-Australian tropics (over 30% in
the noctuid groups studied in the series to date).
There is some variation in proportion of various ecological and geographic
categories amongst the tribes, and data on this have been published by Holloway
& Barlow (1992). The Cassymini show a higher degree of endemism and
restriction to lowland forest than the other major tribes, with endemism lowest
in the Boarmiini. The Boarmiini and Baptini are more strongly represented in
montane habitats.
Genera endemic to Borneo are few. Two are monobasic: Sundasclelia Gen. n.
and Lampadopteryx Warren. Bornealcis Gen. n. has three
species. Some genera such as Peratophyga Warren have a majority of
species in Borneo.
Further collecting in Sumatra and Peninsular Malaysia is likely to reduce the
proportion of species apparently endemic to Borneo. The faunas of the three
areas are typically very similar, and the collecting activities of the
Heterocera Sumatrana Society have reduced the number of Bornean �endemics�
considerably.
Localisation of species within Borneo is uncommon. Two of the Bornealcis species
are restricted to particular mountain ranges, and further examples are found in Milionia
and Dasyboarmia. Abraxas subhyalinata R�ber is only known from Pulo
Laut in southern Borneo and in the Lesser Sundas, a distribution type already
noted for some agaristine noctuids (Part 12). Ruttellerona pulverulenta Warren
is known only from Taganak, a small island off the north coast of Borneo, but
has affinities with taxa in Sulawesi and further east. This is a pattern seen in
some butterflies such as Eurema alitha and Cupha arias that just
reach the northern part of Borneo or occur on small offshore islands (e.g. Pulo
Mangalum), but have ranges extending through the Philippines and further east. Pseudeuchromia
maculifera Felder & Rogenhofer is another ennomine of this geographic
type.
Most genera represented in Borneo exhibit their greatest species richness in
Southeast Asia or the eastern Himalayan region, and attenuate in richness
eastwards into the Australasian tropics. A few, however, are more diverse in the
Australasian tropics and attenuate westwards. Bracca H�bner and Craspedosis
Butler reach their most westerly extremity in Sundaland, but Milionia Walker
extends further onto the Asian mainland, reaching Japan.
One species is adventive. Macaria abydata Guen�e is a Neotropical
species that has established itself widely round the western Pacific margin
during the past decade and a half, probably with Hawaii as a staging post. Its
spread parallels to some extent that of the Leucaena psyllid (See
Macaria abydata Guen�e).
|