|
Cyana
Walker
Type species: detrita Walker,
Bangladesh.
Synonyms:
Bizone Walker (type species perornata Walker); Chionaema Herrich-Schäffer
(type species puella Drury, India); Clerckia Aurivillius (type
species fulvia Linnaeus, �Indonesia�); Doliche Walker (type
species gelida Walker, Bangladesh); Exotrocha Meyrick (type
species liboria Stoll, Sierra Leone); Gnophrioides Heylaerts (type
species flaviplaga Heylaerts, Java); Isine Walker (type species trigutta
Walker, Sierra Leone); Leptothrix Heylaerts (type species tettigonioides
Heylaerts, Sumatra) praeocc.; Macronola Kirby (unnecessary
replacement name for Cyana Walker); Sphragidium Butler (type
species miles Butler, Solomon Islands).
This
genus was known for many years as Chionaema (e.g. Hampson, 1900, who
treated it as an older name than Cyana), but Cyana was later
established as the oldest name (e.g. Roepke, [1946a]; Roesler & K�ppers,
1976). More recently, Cern� (1993) placed the majority of the species in Doliche,
suggesting that the type species of Chionaema was distinguished from
the rest on the grounds of it having a spiny sclerotisation on the aedeagus, and
rather distinctive external features (e.g. grey, irregular fasciation and rather
reduced male secondary sexual characters). However, Cyana
is retained for the group here, following Nielsen et al. (1996), as
the secondary sexual features are certainly within the range of variation seen
in the rest of the genus, the discal area has spots, albeit rather diffuse ones,
and it is possible that the rugosity of the aedeagus is an autapomorphy.
Furthermore, Cern� failed to list any synapomorphies that bring together all the
other species within his concept of Doliche. For example, there is a
complex of grey species (e.g. obscura Hampson, fumea Hampson) in Australia
and New Guinea where there is great variation in secondary sexual features, and
another complex of striking yellow, orange, red and black species (the name Sphragidium
Butler would be applicable to this group) in Australia, New Guinea and the
Melanesian archipelagoes discussed by Holloway (1984). One can agree with Cern�
that there is a fundamental revision required to clarify relationships within
this extensive genus, but for this reason not follow him in prematurely giving
generic status to taxa with unusual morphology but instead seek to identify
distinct groupings within the complex as a first step to elucidating
relationships.
The
presence in the male of some sort of secondary sexual feature between the discal
part of the cell and the costa is perhaps the most consistent feature for the
genus (Fig 10i), and perhaps strong transverse red, orange or yellow fasciation
to the forewings, and spotting in the discal area is also part of the generic
ground plan. The male genitalia have the valves strongly divided, the saccular
process usually ending in a taper and curve to an acute apex. The aedeagus
vesica, when ornamented, tends to have scobinate zones or clumps of spines
rather than single cornuti. The female genitalia frequently have a large
appendix bursae and/or corrugation in the bursa, and the signa, when present,
are scobinate patches typical of the family, one or two in number.
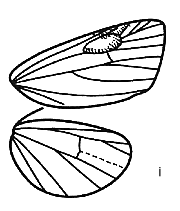
Fig 10i: Cyana pudens Walker
Most
of the Oriental species have typical transverse fasciation on a white ground, as
do many of the African species in the genus. This facies type extends as far
east as the Moluccas (e.g. impunctata Felder), but the rest of the
Australasian species belong the the grey or Sphragidium complexes
mentioned above. Atypical facies is seen also in the dark, narrow-winged
Sundanian species flaviplaga Heylaerts and tettigonioides Heylaerts
(see below). These both have in the aedeagus vesica a long diverticulum that
terminates in a fusion of a few large but very short spines not seen in any
other species examined. They are probably a sister-pair for which the
genus-group name Gnophrioides is available.
Amongst
the typical species there are a number of groups that can be defined on
particular features of facies or male genitalia, as discussed below. The
Philippines appear to have been a centre of speciation for several lineages, as
indicated by Cern�, whereas Sulawesi is characterised more by single,
morphologically isolated endemics. The structure of the androconial patch of the
male forewing may provide some guide to species groupings. Amongst Bornean
species, it usually consists of a circular pad on the underside just basal to
the postmedial. In perornata Walker this is joined by an elliptical pad
basal to it, twice as long with a longitudinal groove. A similar secondary pad,
but without a groove, is seen in conclusa Walker and the malayensis Hampson
group, but is absent or much reduced in cruentata Talbot, maiae sp.
n. and inconclusa Walker. The secondary pad is very much larger than the
circular one in horsfieldi Roepke and costifimbria Walker, both
of which have very much larger fringes of scales from the costa on the upperside
of the forewing. The remaining species lack a secondary pad and have the
circular one much smaller or, in the case of infantula Hampson,
vestigial.
As
well as the works cited above by Roepke and Cern� covering the S.E. Asian fauna,
some Sumatran species were described by Roesler & K�ppers (1976).
The
life histories of three Javanese species not represented in Borneo were
described by Piepers & Snellen (1904). The larvae (all illustrated) are
generally dark, marbled blackish brown or blackish grey, with groups of long
black setae on verrucae. There are often paler whitish, ochreous or red markings
on the thorax, in the anal area and in rows in each segment, often incorporating
the verrucae. A similar larva of C. hamata Walker from Japan was
illustrated by Issiki et al. (1965) and Sugi (1987), the latter also
illustrating the cocoon and pupa.
The
cocoon is much larger than the pupa, ovate, suspended from a vertical surface,
consisting of a very widely spaced but regular net of silk, including the
secondary setae, through which the pupa, suspended vertically in the centre, can
be seen clearly (Fig 3). The secondary setae are set to curve over two apertures
in the outer cocoon, top and bottom.

Fig 3. Cocoon and pupa of Cyana sp.,
Peninsular Malaysia. Scale is 1cm
The
larvae of all three species fed on mosses that grow on damp walls or
tree-trunks. Sugi (1987) also noted that this genus feeds on lichens. Records
from higher plants (e.g. Yunus & Ho (1980); Zhang (1994); unpublished IIE
records) may be erroneous, merely records of the tree on which moss or lichen
was growing. However, Kuroko & Lewvanich (1993) noted larvae of one species
as feeding on leaf tissue (see Cyana sp.
4802).
<<Back
>>Forward <<Return
to Contents page
|