|
Camptoloma Felder
Type species: erythropygum Felder (= interiorata Walker),
Japan, China.
Synonym: Leucopardus Hampson (type species tigrinus Hampson)
syn. n. Taxa in this genus were originally placed in the Arctiidae, and Roepke,
in his description of Leucopardus mirabilis (see below), concurred with
this though indicating the genus was aberrant. Mell (1943) erected the subfamily
Camptolominae for the genus and placed this in the Noctuidae in association with
his concept of the Sarrothripinae (Sarrothripinae and Chloephorinae) because of
similarities of the genitalia and early stages (such as a boat-shaped cocoon).
It was promoted to full family status by Inoue & Sugi (1958-1961) but
relegated to the Arctiinae by Inoue et al. (1982). The history has been
reviewed by Kitching (1984: 208). Holloway (1976) also drew attention to the
chloephorine-like structure of the genitalia of Leucopardus, whilst
listing the genus under Hypsidae: Nyctemerinae.
The genus lacks a tymbal organ, the most useful definitive feature of
the Arctiidae, and, whilst lacking a counter-tympanal hood, has a homologous
sclerite posterior to the first abdominal spiracle, a noctuid feature. This
latter character can be seen best in the female, but in the male the basal
abdominal sternite is considerably modified into a pair of pouches set in a
ventral cleft (Fig. 70), possibly sound-producing organs. A. Watson (pers.
comm.) has examined numerous features of the genus, yet has found only one
potentially arctiid character: two subventral setae (cf. one) the larval
meso- and metathoracic segments, (see also Kitching (1984: 223)), though a full
survey of this feature across the Arctiidae and Noctuidae is needed. It occurs
in larvae of Earias species, placed in the Noctuidae: Chloephorinae.
The basal pouches in the male may also indicate the genus is
chloephorine as similar modification is seen in Pseudoips Hübner, a
typical chloephorine (I .J. Kitching, pers. comm.), and in typical
members of the tribes Hylophilini (=Chloephorini), Careini and Ariolicini
of Mell (1943). Extreme development of this feature is seen in the chloephorine
genus Tympanistes Moore, known to produce a clicking sound or a bat-like
squeak. The structure is also referred to by Kobes (1988). Such modification is
not seen in Westermannia Hübner, so this genus must be excluded from any
concept of the Chloephorinae defined on its presence.
The male genitalia show features reminiscent of the chloephorine noctuid
genus Calymera Moore (see Holloway, 1976): long transtillae that extend
anteriorly into the body cavity when the valves are not spread (fused in Camptoloma
but free in Chloephorinae); the tegumen is expanded on each side at the
junction with the vinculum; there is a subbasal lobe interiorly on the valve
costa; the valve is distally rounded, with a corona of numerous fine setae
directed basad.
These genitalic features and the basal pouches in the male are common to
both Leucopardus and Camptoloma. They also share the following
features that are probably apomorphic and support their synonymy: a strongly
bowed basal section of the forewing costa; wing pattern elements such as a
sequence of oblique black or grey stripes and an orange or yellow patch at the
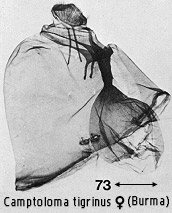
forewing tornus; an extended, rather membranous seventh abdominal segment in the
female, invested with a dense covering of fine scales (Fig. 73), presumably used
to protect the egg-mass.
Other features include filiform male antennae, a rather elongate male
retinaculum, and a rounded, simple bursa copulatrix, lacking signa, with the
ductus seminalis arising from it.
The larva of the type species was described and illustrated by Mutuura et
al. (1965) as follows (from a translation by H. Inoue sent to A. Watson):
Length of body about 35mm. Body cylindrical, little tapering towards
cephalic and caudal ends. Head and caudal tip small. Setae long, their apices
pointed. Body densely covered with minute straight setae, apices of which are
pointed, but they are invisible to the naked eye. Base of each seta flat, SD1 on
each thoracic segment and L2 on prothorax minute, SV on each segment double. SV
on 1st abdominal segment double, those on 2nd segment triple, each of them being
independent, not on accompanying protruberance. SV on 7th and 8th abdominal
segment single. SD1 on 8th segment placed just above spiracle. Setae at side of
each proleg are three, secondary setae not developed. SD1, 2 on prothorax are
placed below shield. Among crochets of prolegs those at the central one-third
are long, becoming shorter towards both sides. Head roundish, densely covered
with minute wrinkles. Forehead small, longitudinally long, vertex roundish.
Coronal suture about twice length of forehead. Ad-forehead is broad throughout
its total length. 1st and 2nd ocellus approximated, distances of 2nd, 3rd and
4th are nearly the diameter of eye. Mandible with three thick teeth; there is
an internal tooth in a diamond-shaped protruberance at the bottom of the inner
side. Spinneret thick and short, about three times length of width at the
middle. Two setae from below the anus are curved, forming anal fork.
Head black, two thick whitish stripes at side.
Frontoclypeal area and base of antennae greyish white. Body blackish brown, with
narrow yellowish red stripes. The dorsal line, three subdorsal lines, two
supraspiracular lines and a subspiracular line slender, minutely wavy.
Prothoracic shield black, with greyish white dorsal line. Anal plate black,
margined with greyish white. Spiracles yellowish brown, ringed with brown.
Thoracic legs black.
Larvae gregarious, making a common pouch-like nest.
Hibernates in a young larval stage, again active around April and full grown
during May and June. The silken nest is greyish white, attached to a tree-trunk
or twigs. A large one measures 9cm in diameter. Larvae stay in the nest during
daytime and come out to eat leaves at night. The last instar larvae crowd on a
tree trunk even during daytime. When fully-grown, they walk down to the ground
and spin thick yellowish white cocoons between fallen leaves. Adults fly in
June.
Recorded host-plants are all from the Fagaceae (Mell, 1943; Mutuura et
al., 1965): Quercus, Castanopsis, Castanea, Lithocarpus.
The genus will be examined further in conjunction with the
Sarrothripinae and Chloephorinae when that part of the series is undertaken.
<<Back
>>Forward <<Return
to Contents page
|