|
When
this series was originally planned, the intention was to publish the treatments
of the traditional (e.g. Hampson, 1902; Nye, 1975) concepts of the Catocalinae
and Ophiderinae in two parts, the first containing the Catocalinae and some
well-defined groups of the Ophiderinae, and the second containing the rest of
the Ophiderinae. The traditional means of separating these two groups was the
presence of spines on the mid-tibia (Catocalinae) or not (Ophiderinae), an
unsatisfactory criterion as discussed on p. 11 and in a detailed historical
review of the classification of the group by K�hne & Speidel (2004).
However,
since that original intention, the classification of the major groupings of the
�quadrifines� (with a strong vein M2 in the hindwing) has gone progressively
into meltdown. This was commenced by a major and comprehensive review of
noctuid classification by Kitching (1984: fig 4), who presented a cladogram
representing relationships between the various noctuid subgroups. He united the
Catocalinae and Ophiderinae in one group, the Catocalinae, that he considered
was �better characterised [than the Hypenodinae and Hypeninae] (fused pleural
sclerite in the male genitalia and pupa with a whitish bloom) but the majority
of genera and species of this vast subfamily have yet to be examined,
especially with regard to the pupal character�. He showed three branches in the
cladogram, though he indicated these were merely examples of many genus-groups
that could be recognised. Nevertheless, one of these included Catocala Schrank and Othreis Hübner (with Ophideres Boisduval a synonym),
the type genera of the two original subfamilies, united by possession of a
chitinous projection from the inner margin of the tympanal frame.
The
situation was reviewed further by Speidel & Naumann (1995a) who suggested
that the monophyly of the subfamily was highly doubtful and recommended as a
first step the definition of monophyletic groups within the complex and only
then to attempt to establish the phylogenetic relations for these monophyla.
This philosophy has been mostly followed subsequently, e.g. by Speidel et al. (1996a),
Kitching & Rawlins (1998) and Holloway et al. (2001). It was
also endorsed at a workshop on Noctuidae classification at the Societas
Europaea Lepidopterologica (SEL) Congress at Kors�r, Denmark,
in June, 2002, and also by K�hne & Speidel, 2004. This was attended by
myself and most of the authors cited above. At the meeting it was suggested
that a review of family-group names would provide a useful foundation for the
eventual nomenclature of the group, and this
one has just been published (K�hne & Speidel, 2004; Speidel & Naumann,
2005); these total well over the 70 encountered during a rough survey of the
literature in preparation for this volume, and many are noted also by Kitching
(1984). As well as the dissections of male and female genitalia undertaken for
the genera in the Bornean fauna to complement those already available in BMNH,
a similar programme of dissection was undertaken for these family-group names.
This resulted in a collection of about 2000 slides. These were also made
available for the work of Fibiger (2003) on the European Catocalinae during
visits of that author to London,
when valuable discussions on the classification of the group, following those
in Kors�r, proved very fruitful also to developing the approach to the
classification adopted here.
However,
prior to the SEL meeting in 2002, Speidel et al. (1996b)
suggested a division of the Catocalinae/Ophiderinae complex into two groups,
Catocalini and Calpini, that reflected proboscis structure, and placed the
Erebini within their Catocalinae. They did not examine closely many of the
genera in the �framed corematous� sequence discussed below. They noted that the
name Erebini is older than Catocalini; both are older than Calpini. They
suggested that an application to the ICZN should be made to conserve the name
Catocalinae for the subfamily as a whole; K�hne & Speidel (2004) proposed
giving Catocalinae precedence under ICZN article 23.9.1. Later, Fibiger (2003)
placed the Erebini in his concept of the Calpinae (see below), so, if his
classification is accepted, Erebini would have priority for this group also;
see also the arrangement of Forbes (1954), reviewed by Kitching (1984: 201).
Kitching
& Rawlins (1998) divided the complex into Catocalinae, (including ) and a
restricted concept of Calpinae (including Gonopterini and Anomiini, here
treated as Scoliopteryginae), noting the unsatisfactory nature of the
classification, including paraphyly of their Catocalinae, and recommending the
approach endorsed at Kors�r.
Yela
& Kitching (1999) recognised the Catocalinae, Calpinae and Scoliopteryginae
(as Gonopterinae) as subfamilies, and suggested that the second and third were
more closely related to each other than to the Catocalinae. They excluded the
Bagisarinae, Aediinae and Tytinae, indicating these are probably best included
in the trifine sequence of subfamilies (see p. 26).
Fibiger
(2003), whilst attempting to define monophyletic units within the European
fauna, nevertheless felt it necessary to use and define various systematic
levels or ranks: tribes, subtribes, genus-groups, subgenera and species groups.
He also attempted to divide a much wider selection of genera into the two
subfamilies recognised by
Kitching & Rawlins (1998): Catocalinae and Calpinae. As will be seen from
the genera he listed, very many are those occurring in Borneo, and his
assignments of these are noted in the section following to enable the reader to
make cross-reference more easily.
For
various reasons which will be detailed in the sections following, it has not
been possible to follow the system of Fibiger (2003) in its entirety. This
system was presented as a highly structured phylogeny, at least for the
Catocalinae. In fact I have reverted to the spirit of the discussion at
Kors�r,
endorsed by K�hne & Speidel (2004), and merely identified tribes,
interspersed by the approach of Forbes (1954) who recognised sequences of
miscellaneous genera. I do not have sufficient confidence in my interpretation
of the value of various characters to impose a more rigid hierarchy on the
group, but hope that I may have provided some pointers that may enable others
to do this. The dangers of adopting too precise an approach using only a
limited suite of characters is illustrated by the fact that those used by
Fibiger (2003) have caused him in a few cases to divide between his Catocalinae
and Calpinae groups of genera clearly linked by synapomorphies (e.g. as
discussed below and on pp. 122 (Tinoliini), 160 (Erebini) and 275 (Anobini)).
Nevertheless,
I have tried to sequence the groups from what might be termed �core
catocalines�, that are perhaps more closely allied to Catocala, through to a
sequence of genera that may represent a more paraphyletic tail to the group and
which are referred to as the �framed corematous� sequence, sharing particular
features of the male eighth abdominal segment that also occur more widely in
the Noctuidae as described on pp. 14-16.
This is
to some extent a compromise between the concepts of Kitching & Rawlins
(1998) and Yela & Kitching (1999) on the one hand and Fibiger (2003) on the
other, retaining the concepts of the Calpinae and Scoliopteryginae of the
former and the core catocalines of the latter more informally, and recognising
the pervasive paraphyly of the complex as a whole.
These
core catocalines show some considerable overlap with the concept of the
Catocalinae outlined by Fibiger (2003), as does the recognition of some of the
groupings within them. In the sections following, several of characters are
highlighted that are shared fairly widely amongst these groupings, two of which
were noted by Fibiger (2003): the structure of the juxta in the male genitalia
(p. 18); and that of the seventh segment and the position of the ostium
relative to it in the female genitalia (p. 20). Fibiger (2003) has made a
detailed study of the latter character and its importance, and has adopted a
scoring system for the position of the ostium relative to the seventh and
eighth segments. The survey of Bornean genera supports his assertion of its
importance. The feature of the juxta appears to be more widespread, but it is
enhanced and developed in the core catocalines, where it is usually fused with
the anellus (Fibiger, 2003).
The
�framed corematous� condition of the male eighth segment (p. 14) is probably a
plesiomorphy within the Catocalinae, hence the unmodified state of this
segment, with short, widely separated and more or less parallel apodemes on the
tergite, may represent another apomorphy for the core catocalines. However, the
plesiomorphic condition is seen in a somewhat vestigial form in some species of Catocala
(p.
38)!
Features
of the proboscis or tongue may also be significant, as suggested by Speidel et al. (1996b) and reviewed
on p. 10, perhaps helping delimit the core catocalines and also a group around
the Calpini and Scoliopterygini, tribes which initiate the �framed corematous�
sequence of genera on p. 213.
Fibiger
(2003) did not include characters of early stages in his analysis, but the
�ophiusine� (Bell, MS) type of larva (p. 22) with a pair of dorsolateral
tubercles on A8, and presence of a waxy or powdery bloom on the pupa (p. 23)
may also facilitate recognition of core catocaline groups, though the latter
character occurs in a few of the �framed corematous� genera and (Kitching &
Rawlins, 1998) in some Herminiinae.
The
molecular analyses of Weller et al. (1994) included
a core catocaline, (Catocala), a core calpine (Plusiodonta Guenée) and a
hypenine (Plathypena Grote) from the quadrifine sequence of subfamilies
as well as a sample of six trifine genera. Whilst the latter were combined when
mitochondrial and nuclear DNA data were grouped together, the three quadrifines
were intermingled within the seven Arctiidae included.
The
molecular analyses of Mitchell (1998), Fang et al. (2000) and
Mitchell et al. (2000) only included three taxa from the Catocalinae /
Ophiderinae complex. Two were core catocalines, a species of Catocala and one of Caenurgina McDunnough from
the Euclidiini. They selected the type species of Hypsoropha Hübner as a
representative of the Calpinae, also included in his concept of this group by
Fibiger (2003). This is not related to the Calpini as circumscribed here, but
has a number of unusual features that make its placement unclear. The male
eighth segment is distinctive, the sternite modified into two narrow processes,
and the tergite broad, without obvious apodemes. The juxta is weak and also
atypical. The female has the ostium intimately associated with the eighth
segment. The three taxa form a clade in an analysis combining data sets from
two different genes (Mitchell et al., 2000), which
generated a stronger phylogenetic signal than analyses of each gene separately,
though one of the latter also grouped the three, the other grouping Hypsoropha with the
Aganainae. The analyses reported by Fang et al. (2000) using the
dopa decarboxylase gene give no grouping of the three in a clade, or grouped Caenurgina with Hypsoropha, excluding Catocala.
As with
the analyses of Weller et al. (1994), quadrifine examples are grouped
with the Arctiidae and Lymantriidae rather than with the trifine group of
examples in analyses with combined DNA data. These results raise serious
questions about the status of the Noctuidae, indicating that the family is not
monophyletic.
The
order of groups in the systematic section can be related to the treatment of
the European fauna by Fibiger (2003). For reasons given above, the tribes
Catephiini, Pericymini and Pandesmini are treated separately from the Ophiusini
rather than united with them. The groups up to the Ommatophorini overlap
completely with his Catocalinae. The Hulodini, a tentative assemblage, include
two genera, Hulodes Guenée and Ericeia Walker, from his
Calpinae with two from his Catocalinae. The Tinoliini include one genus from
each, as do the Erebini after the first miscellaneous series of genera. This
miscellaneous series includes genera where the male eighth segment appears to
be of the catocaline type and includes seven further genera listed as
catocaline by Fibiger (Erygia Guenée, Anisoneura Guenée, Platyja Hübner, Sympis Guenée, Bamra Moore, Dordura Moore, Varicosia Hampson), as
well as several that are not. The second sequence of miscellaneous genera
commencing on p. 183, where the male eighth segment and other features are
unusual in some way, includes two more genera listed by Fibiger as catocaline: Batracharta Walker (Phorica Walker is
related but not included by Fibiger, as a female had not been dissected prior
to his study of the slides in London) and Bocula Guenée. His
catocaline list also includes a few more genera from the �framed corematous�
sequence of this work: Blasticorhinus Butler, Delgamma Moore, Dinumma Walker, Gesonia Walker, Hamodes Guenée, Homodes Guenée, Loxioda Warren, Plecoptera Guenée and Tephriopis Hampson. Dinumma is here placed
tentatively with the Scoliopterygini but has a bloom on the pupa (p. 242). Tephriopis, and probably Plecoptera, are allied to Anoba Walker, placed
in the Calpinae by Fibiger, but are here grouped together in the Anobini (p.
275). Delgamma and Hamodes, with Hypospila Guenée, a
Fibiger calpine, are all treated in the third sequence of miscellaneous genera
starting on p. 264 that includes species with similar facies and a framed
corematous male eighth sternite combined, in the three genera just noted, with
a pupal bloom. Homodes is a very distinctive genus, particularly in the
characteristics of the larva, but may have a pupal bloom and is therefore also
included in the third sequence (p. 272). Blasticorhinus and Loxioda may be related
(see p. 429). Gesonia has rather ambiguous features (p. 426) but has been
included here in the �framed corematous� sequence of groups. These last three
genera are all treated here in the sixth miscellaneous sequence.
The
third miscellaneous sequence comes after treatment of the Scoliopterygini,
Calpini and Phyllodini, and marks the beginning of a series of groups that are
drawn mostly from the old concept of the Ophiderinae and placed by Fibiger
(2003) in his broad Calpinae grouping. Most have a framed corematous male
eighth abdominal segment or a variation on that theme. The third sequence is
followed by the Anobini and then by the fourth miscellaneous sequence, an
otherwise heterogenerous group of genera that share facies and build with the
Scoliopterygini but lack the definitive features of that group. The Episparis Walker group of
genera that follows is defined primarily on features of the female ovipositor,
but contains genera where the wings are angled in the centre of the distal
margin as in the fourth miscellaneous sequence and in the fifth that follows.
The fifth starts with genera that share facies features with the Episparis group and grade
to genera such as Mecodina Guenée that share facies features with the tribe
Pangraptini that follows. The Bornean Pangraptini consist entirely of the genus
Pangrapta
Hübner,
but no features were located that could enable the concept to broaden to
include the previous two groups. The three groups together include a number of
larval host records from the plant family Rubiaceae (p. 32).
The two
large groups that follow are more clearly defined, the Saroba Walker group
defined primarily on abdominal features and the Boletobiini to some extent on
these but also on the loss of the two anterior pairs of prolegs on the larva
and a fungus-feeding biology.
The
treatment concludes with a sixth miscellaneous sequence of genera. Many of
these have characters that suggest they may be better classified in other
quadrifine subfamilies such as Hypeninae and Herminiinae, and will be reviewed
further with these and with the Acontiinae, Eustrotiinae and Eublemmini complex
in the two parts of this series that will cover the remaining Noctuidae.
The
Catocalinae / Ophiderinae complex is of considerable economic importance as it
contains many agricultural pests. These include both defoliating larvae and
fruit-piercing adults, occasionally both within the same genus or species. The
defoliators affect both tree and field crops, and some can have impact on
forestry plantations. Fruit-piercing is dealt with in more detail on p. 32; it
involves many core Catocalinae but also the Calpini and Scoliopterygini, and is
intimately related to modifications of the tongue (p. 10). Amongst the Calpini,
the genus Calyptra Ochsenheimer includes several species where this
tongue modification has facilitated the piercing of mammalian skin and
blood-sucking (p. 32).
Many
species in the group are highly mobile and have very wide geographical ranges;
some migrate over long distances, and species of Mocis Hübner, the Anomis Hübner group and
Anticarsia
Hübner
have proved particularly successful in reaching remote oceanic islands
(Holloway & Nielsen, 1999). Of 30 noctuid species recorded as migrant to
Norfolk I., half are Catocalinae in the broad sense (Holloway, 1996). Several
species, particularly in the Ophiusini, are characteristic of open habitats in
Borneo. These biogeographic and ecological features are discussed further on p.
33.
Larval
host-plant specialism is reviewed on p. 31. A high number of genera and wider
groups show specialism to Leguminosae, but other plant groups with genera
specialist on, or sharing strong preference for them include Acanthaceae, Ebenaceae,
Euphorbiaceae, Gramineae, Malvales, Menispermaceae, Palmae and Sapindaceae. One
group, the Boletobiini, has larvae that feed on bracket fungi (pp. 32 and 372).
Adult
features
The
head
The
head offers one feature that may be reliable for identifying Catocalinae in the
broadest sense: lack of scales on the lower clypeofrons, immediately above the
tongue, as discussed in detail below (p. 24), where it is used in combination
with other characters to exclude a number of genera traditionally placed as
catocalines or ophiderines.
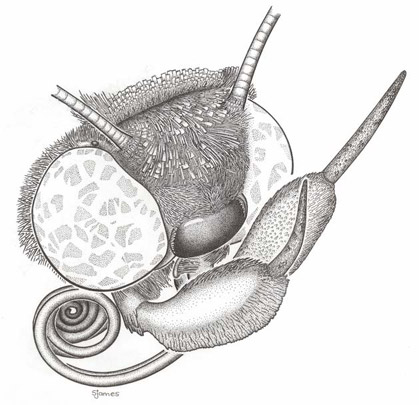
Fig 1.
Head of
Ugia viridior
sp. n., showing the unscaled
clypeofrons and typical catocaline labial palps. Artwork by Shayleen James.
This
area is usually obscured by the labial palps which are characteristically well
developed, ascending to vertical, with a third segment that is approximately as
long as the second but very much more slender. Both these and the scaleless
clypeofrons are illustrated in Fig 1. This general structure is seen also in
the Aganainae (Holloway, 1988: 77) and Nolidae (Holloway, 2003: 194, 220). The
third segment is particularly long, scaled and prominent, and the palpi more
porrect (directed forwards) in Schistorhynx Hampson, Brontypena Holland, Lignicida Swinhoe and Aburina M�schler, and
palps are generally long and slender in a few genera such as Stenocarsia Hampson, Hyposemansis Hampson, Pangrapta Hübner and Egnasia Walker. Several
genera have the third segment much shorter than the second, most of these
outside the �core catocalines� treated up to p. 130, and some being rather
distinctive genera such as Bocula, Hypocala Guenée and Homodes. In a few
genera (e.g. Chilkasa Swinhoe) the third segment is downturned.
The
male antennae are usually fasciculate or ciliated. Bipectinate antennae are
much rarer but occur in several of the genera of the Scoliopterygini (p. 213),
of the Episparis Walker group of genera (p. 292), some species in the
Sypnini, and within Avatha Walker. Other genera where they occur include, with
the antennae very long in the first three, Ugia Walker, Heterospila Guenée, Tochara Moore, Pseudosphetta Hampson and Crithote Walker. In Platyja Hübner they are
bipectinate with an additional ventral serration, giving them a tripectinate
appearance (Zilli, 2001; p. 141). The male antennae are noded in Lutzugia Gen. n. and swollen in Psimada Walker and Pericyma
cruegeri Butler.
Many of
the genera included in the Catocalinae, particularly those where the moths are
relatively robust, have been noted to feed as adults on fruit and, in one
genus, on lachrymal secretions and piercing mammalian skin to feed on blood.
Whilst several other families of moth have ben recorded as lachrymal and
carrion feeders (reviewed by Holloway et al., 2001), fruit
feeding has been observed predominantly in the Catocalinae. These habits are
associated with modification of the proboscis or tongue; biological aspects are
reviewed in more detail on p. 32, but corresponding morphological features are
discussed here.
The
most comprehensive review of tongue structure is that of Speidel et al. (1996b), but
other publications of particular relevance are those by B�ttiker (1962),
Bänziger (1982), Wu & Chou (1985) and B�ttiker et al. (1996). Speidel et al. recognised differences
between the more basal and the distal part of the tongue, the latter having
numerous styloconic sensilla, and the former having a few chaetiform sensilla.
Both regions were found to show structural diversity and to be of relevance to
the classification. The basal zone is much more extensive than the distal one,
but several of the categories Speidel et al. recognised occur
both within and outside the Catocalinae. The categories, with Bornean genera
noted, are as follows:
|
Smooth, simple.
|
Pericyma Herrich-Schäffer, Scoliopterygini (Scoliopteryx Germar), Eudocima Billberg.
|
|
Smooth, lamellate.
|
Bematha Walker, Oxyodes Guenée, Calesia Guenée (part);
and Aganainae, Herminiinae.
|
|
Smooth, with dentated ridges.
|
Boletobiini (Parascotia Hübner), ?Pangraptini
(Zethes
Rambur);
in several non-catocaline groups, particularly the Euteliinae and
Stictopterinae.
|
|
Smooth, with cuticular microprojections.
|
Very basally in Calyptra Ochsenheimer, more
strongly in Toxocampini (Lygephilini); widely in trifine subfamilies and also
in Nolidae.
|
|
Fluted, simple.
|
Catocalini, Erebini, Calesia (part).
|
The
smooth and fluted conditions refer to the semicircular ribs of the proboscis,
the latter having the ribs broken by longitudinal depressions. The Eublemmini (Eublemma Hübner) also
share this condition. No clear signal is apparent from these data; the sample
of taxa was modest, and a wider survey will be necessary to assess what pattern
there is. However, the Catocalinae as covered here show a tendency for
simplicity.
There
is a stronger phylogenetic signal from the distal part of the tongue, where
some of the Catocalinae have a smooth apex to the proboscis; the nodulose
condition is seen widely in the noctuids, particularly in trifine subfamilies
and also in other noctuoid families, and was therefore considered by Speidel et al. (1996b) to be
the plesiomorphic condition. A smooth apex is found in the Plusiinae, the
acontiine genus Metaemene Hampson (with a Bornean type species, atrigutta Walker,
illustrated by Barlow (1982)), and in the following Bornean catocaline tribes
or genera: Catocalini, Pericymini, Erebini, Calpini, Scoliopterygini. Within
this last group, the Calpini and Scoliopterygini have the apex of the tongue
furthermodified by strongly sclerotised, erectile, reversed hooks, considered
by most authors cited above to be modified sensilla styloconica. Strong,
enlarged spines have also been noted in Serrodes Guenée
(B�ttiker, 1962; Bänziger, 1982), the Ercheiini, Pericymini and Phyllodini, and
the ophiusine genera Artena Walker, Ophiusa Ochsenheimer and
Thyas
Hübner,
but only in Pericymini are these definitely erectile (Bänziger, 1982).
Speidel et
al. (1996b) considered the occurrence of a smooth apex in Plusiinae
and Catocalinae to be homoplasious. If it is synapomorphic within Catocalinae,
it embraces representatives of both Catocalinae and Calpinae as recognised by
Fibiger (2003), and would suggest that this division requires reassessment.
Speidel et al. noted that the Plusiinae have few sensilla
styloconica. In many catocalines they are situated on the dorsal part of the
tongue, observed in Catocalini, Erebini, Hypopyrini and several Ophiusini (see
also Wu & Chou (1985)), but not in Calpini and Scoliopterygini. In these
former groups the sensilla are modified into blade-like structures that are
also seen in other fruit or lachrymal feeding groups (B�ttiker et al., 1996).
A
feature that is shared by at least some Calpini and Scoliopterygini is an
additional type of sensillum that is apically furcate (Speidel et al. 1996b). Speidel et al. also noted that
the Phyllodini had been associated with these tribes by Berio (1959), but had
not been able to explore this further.
The
legs and wings
The
legs provided the traditional means of sorting most of the quadrifine noctuids
into Catocalinae and Ophiderinae, namely whether the mid-tibia in particular
was spined or unspined (Hampson, 1902). This was challenged to some extent by
Hampson himself (Kitching, 1984), but particularly by Prout (1921), Richards
(1932) and Berio (1959, 1992), all noting that both states could occur within
otherwise closely related taxa, several examples of which can be found in the
Bornean fauna, e.g. in Avatha, Buzara Walker, the
Sypnini (see Berio & Fletcher (1958)), the Calpini and Phyllodini (pp.
246-262) and in Arcte Kollar (p. 28). K�hne & Speidel (2004) have provided a
detailed review of literature on this feature. Nevertheless, occurrence of the
feature is concentrated within the sequence of taxa referred to the core
catocalines on p. 5-6, is general to tribes such as the Catocalini, Ercheiini,
Pericymini, Erebini and Hypopyrini and predominates in the Ophiusini.
Berio
(1992) attempted a fresh segregation of groups within the complex, again using
mainly characters of leg structure but trying to accommodate variation within
genera, between sexes (he suggested spining tended to be more developed in
females), and between legs. He recognised eight categories on this basis,
grading from spiny to smooth, and organised the genera studied into tribes and
subtribes on these grades and in relation to general similarities in other
features. The three tribes were: Catocalini (solo spinose: spined only);
Ophiderini (miste: mixed); Phytometrini (inermi: unarmed,
smooth). He drew particularly on the work of Wiltshire (1990) for tribal
nomenclature, though several of the names are unavailable according to the ICZN
Code (K�hne & Speidel, 2004; Speidel & Naumann, 2005). The Catocalini
included the Catocalini, Ophiusini and Ercheiini as presented here, and the
Ophiderini included the Erebini and Calpini as well as the Arcte group (see p.
28). The Phytometrini included the Scoliopterygini, but also the Catephiini
(including Aediinae; see p. 84) and Ericeini (here placed in the Hulodini). The
Pandesmini and Toxocampini as presented here were divided between the second
and third of his groups.
Speidel et
al. (1996a) suggested that a putative autapomorphy for the
Catocalinae sensu lato is presence of a mid-tibial corema. Kitching &
Rawlins (1998) noted this as a groove containing a brush organ. The males of
very many genera will be noted in the descriptions in the main text to have
their legs variously modified by hair pencils and scale crests, often very
prominently. It was beyond the scope of this survey to assess these in detail,
as it would require removal and microscopic examination, including descaling,
of all legs on one side in each case. However, it is undoubtedly a task that
must be undertaken to improve our understanding of the classification of the
Noctuidae, and will no doubt revisit the work of Berio reviewed in the previous
paragraph. Degree of development of hair pencils and coremata on the legs should
be contrasted with development of posterior abdominal brush organs as part of
the �framed corematous� condition of the male eighth segment (p. 14).
Structures on the hindlegs might be expected to interact with these, whereas
those on the other pairs of legs may usurp their function and be correlated
with their reduction.
The
wing venation has not been studied in detail for this work, though some genera
have it modified. The hindwing is quadrifine with M2 present, strong, and
usually arising from a point adjacent to M3. Weakness and loss of M2 was taken
to characterise the trifine subfamilies or higher Noctuidae, though some of
those currently placed in this trifine sequence (e.g. by Speidel et al. (1996a)), such
as the Aediinae, Plusiinae, Eustrotiinae, Bagisarinae and Acontiinae, are in
fact quadrifine. Again, as with the spined mid-tibia characters discussed
above, this has led on occasions to otherwise closely related taxa being
separated widely in the classification. This is illustrated in particular by
the new concept of the genus Chorsia Walker (p. 405)
that brings together species previously in the Amphipyrinae, Acontiinae and
Ophiderinae.
Fibiger
(2003) identified a wing pattern feature that, from a cladistic analysis he
performed by hand, he considered to be one of the three most reliable for
identifying a Catocalinae core group of tribes and genera. This is a black or
darker terminal patch or fringes on the hindwing between veins M3 and CuA1.
This character is sometimes lost where pattern is strongly modified, but
Fibiger (2003) suggested that, even in such groups, there were member taxa that
retain this patch to some extent. Interpretation of this character has been
difficult in the much more diverse Bornean fauna, and it is possible that the feature
represents more of a rupture of a generally dark hindwing ground by distinctly
paler sections of the margin on each side. Its occurrence is far from general,
but can be observed in the Catocalini generally, in a few Ophiusini such as Artena Walker, Grammodes Guenée and Achaea Hübner, in the
Ercheiini, in some Hypocala Guenée, Fodina Guenée and in
the Catephiini. It also occurs in the Aediinae, Tytinae and Armadini,
considered by Fibiger (2003) to be catocaline, but the first two are possibly
better associated with the trifine subfamilies as discussed on p. 26. Pale
sections of hindwing margin with a wider dark section in between are seen in Serrodes Guenée and,
anteriorly, in some members of the Sypnini. In many Eudocima the margin is broken by a series of pale
patches or is entirely pale. The feature in its many guises appears to be
associated strongly with paler flash coloration (a basal zone or prominent
spotting), particularly that involving white or yellow, but also pale red in Catocala. Occurring as
it does in Hypocala and Fodina, both placed by
Fibiger (2003) in his Calpinae, in the Aediinae and, in a more broken form,
even in some Agaristinae, it may possibly represent a homoplasious feature
associated with the development of at least a partial diurnal habit (e.g. for
escape on disturbance) in the more open habitats of savannah in the tropics and
semi-arid regions in temperate latitudes, rather than an autapomorphy for a
core concept for the Catocalinae.
A
hindwing feature that appears sporadically through the Catocalinae /
Ophiderinae is the presence of a slight obtuse angle on the margin at vein
CuA2. It is seen, for example, in some Ophiusini, in Ugia Walker, in many
of the Saroba group of genera (particularly Trichoblemma Hampson) and
even in some members of the sixth miscellaneous sequence such as Rema Swinhoe and Marapana Moore.
There
is a lot of variation within the broader group in the relation of the hindwing
pattern to that of the forewing. Perhaps the majority of genera has the
hindwing distinctly different to that of the forewing, usually plainer, but
possibly also involving the flash coloration just discussed. But in tribal
groups such as the Pericymini, Hulodini, Amphigoniini, Erebini and Hypopyrini,
the patterning of the hindwing reflects that of the forewing to a greater or
lesser extent, though is always simpler antemedially. There is often a
prominent fascia that is continued from the forewing to an element of the
hindwing, e.g. in genera such as Ugia Walker, Armana Swinhoe and Tochara Moore. The
occurrence of hindwing-forewing similarity becomes perhaps more frequent
amongst the more delicate genera with the �framed corematous� type of male
eighth abdominal segment (p. 14), such as in the Boletobiini and the Episparis Walker and Saroba Walker groups of
genera.
The
variation is probably correlated with resting posture. Groups with hindwing
flash coloration and cryptic, leaf-like forewings such as many Ophiusini and
Calpini tend to have a tectiform (rooflike) resting posture with the hindwings
concealed. Groups with hindwings patterned as in the forewings, both usually
cryptic, tend to have a flatter resting posture with the hindwings exposed as
described for the Erebini on p. 165. Those with a discal ocellate mark on the
forewing such as in the Erebini, Hypopyrini and Speiredonia Hübner mostly
fall into this latter category, though Ommatophora Guenée may be
intermediate. The two genera assigned here to the Pandesmini (p. 101) fall one
into each of these categories, but are related through early stages and genital
features as well as sharing similar larval biology.
Holloway et
al. (2001) suggested that a strong looping of the forewing
postmedial was another facies feature that might be of phylogenetic
significance. However, the exploration of other characters in the course of
this work has indicated that it might be somewhat homoplasious. Also, it can be
obscured and dominated by other pattern developments. The postmedial, when
looped, runs round the discal area from the costa, and then curves round
posterior to the reniform to run basad parallel to the costa, or even curving
back towards it, before angling sharply back to run more or less straight to
the dorsum in a much more basal position. This feature is seen most clearly in
the Anomis Hübner group of the Scoliopterygini, but also in Erygia Guenée, Ommatophora, Speiredonia and possibly Spirama Guenée, three of
these being genera with forewing discal ocelli. It occurs weakly in Hypocala, Hyperlopha Hampson and Polydesma Guenée. The most
extreme development of the loop is in Platyja Hübner as
described on p. 141.
Another
forewing feature that occurs in a number of apparently widely separated genera
or groups, and therefore possibly homoplasious, is the development of a
distinct, usually darker mark, triangular or trapezoidal, on the forewing
costa, defined by the postmedial and submarginal basally and distally and by a
line usually along vein M1 posteriorly. It is seen most extensively and
typically in the Parallelia Hübner group of genera (Holloway & Miller,
2003), but also in Ommatophora, Dordura Moore and in
several instances in the �framed corematous� sequence (see below) of genera;
e.g. Claterna Walker, Leptocola Gen. n., in the
Pangraptini, in taxa in the fifth miscellaneous sequence and in the Saroba group.
The
forewing reniform stigma is also very variable and often distinctive, but
tending in the �framed corematous� sequence to be somewhat bilobed, often with
a dark dot at the centre of each lobe. The discal mark of the hindwing may also
be bipunctate. These features are difficult to score and may be variable within
a single genus, e.g. Tamba Walker. Bipunctate discal markings also occur in genera such as Arasada Moore and Oruza Walker in the
traditional Acontiinae / Eustrotiinae complex.
The
male eighth and basal abdominal segments
The
eighth segment of the male abdomen is often strongly modified in genera of the
Catocalinae / Ophiderinae complex, but such modification is only slight in the
core catocalines, usually restricted to short, broad, widely separated apodemes
on the tergite, though Catocala is somewhat exceptional in this, with
vestigial modification of the type described below seen in some species (p.
38). Given the wide distribution through the noctuids of what is termed the
�framed corematous� condition below, this unmodified state could be apomorphic
and provide support for inclusion of tribes and genera within the core
catocalines. However, the eighth segment is unmodified in the first
miscellaneous sequence of genera from Fodina Guenée to Cryptastria Hampson, and
these genera do not have many other features that are typical of core
catocalines. It is also unmodified in Hypena Schrank, but is
modified in another core hypenine, Harita Moore, and in Ricla Walker (Lödl,
2000).
The
�framed corematous� condition (see also Birch, 1972: 203-4; Poole, 1995: 16;
Lödl, 2000) is found scattered through the noctuid classification. It occurs in
many trifine genera discussed and illustrated by Holloway (1989: 60-61), e.g. Dipterygina Sugi (fig 21), Thalatha Walker (fig 24),
Athetis
Hübner
(fig 38) and, possibly somewhat modified, in the Mythimnini (fig 23, and see
also Zilli & di Giulio (1996)), thus involving several of the trifine
subfamilies. Kitching (1987), in his study of the Plusiinae, noted the framed
corematous condition as occurring in the Stictopterinae and Euteliinae as well
as in the Plusiinae, becoming even more elaborate in some (see also Holloway,
1985). He interpreted simple quadrate sclerites on A8 as plesiomorphic. It is
also seen in the Hypeninae (Lödl, 2000) in genera of the Acidon Hampson / Hiaspis Walker complex
(Lödl, 1998: figs 70, 82) and in Mecistoptera Hampson, a genus
of similar build and facies to Hiaspis but with a prespiracular
counter-tympanal hood and therefore associated with the Herminiinae by Lödl
(1997: fig 5) in his revision; see also Hepatica Staudinger on p.
452. It occurs in some of the taxa discussed on pp. 28-29. Type genera of
family groups within the old Hampsonian concept of the Acontiinae or
Eustrotiinae (Nye, 1975) show varying development of the features. In Eustrotia Hübner the
tergite could be of the framed corematous type but the sternite is atypical,
entire. Acontia Ochsenheimer is more typical, with the coremata
strongly developed though narrow and well separated. Eublemma Hübner has the
tergite relatively broad; though the corematous zone is reduced on the
sternite, lateral rods are present. Other quadrifine groups showing the feature
include Rivula Guenée, Hypenodes Doubleday and Herminia Latreille,
though Lödl (2000) stated that the feature was generally absent in typical
Herminiinae (e.g. as treated by Owada (1987)). The first two genera are
typical; Herminia has the sternite elongate with long lateral rods,
though the tergite is atypical, reduced, with long, slender, separated and
unsplayed apodemes.
The
occurrence of this feature outside the Noctuidae needs further study, but where
coremata are present in the Lithosiinae (Holloway, 2001: figs 268, 270, 272,
316) and Arctiinae: Spilosoma (Holloway, 1988: 40), the structure of both sternite
and tergite is different. Kitching & Rawlins (1998) illustrated the
structure in the arctiid subfamily Ctenuchinae and noted that such organs can
be present on almost any abdominal segment, but most often on the eighth. A
diversity of structure of the eighth segment is seen also in Nolidae (Holloway,
2003), but this does not appear to involve coremata. The structure might be a
ground plan feature for the Noctuidae (but also including the Euteliinae +
Stictopterinae), though molecular evidence (Mitchell et al., 2000)
indicates that the noctuids are paraphyletic relative to the Arctiidae and
Lymantriidae.
Both
tergite and sternite are modified in the �framed corematous� condition. The
sternite typically has the lateral and anterior margins sclerotised only
narrowly, forming a three-sided frame for a membranous anterior lacuna that is
frequently developed into a pair of coremata or, more rarely, one central one.
Lödl (2000) referred to this sternal structure as the posterior abdominal
brush. The distal margin is less distinctly sclerotised but usually supports a
band of deciduous scales or hairs that are represented by their bases in a
slide preparation. The �sides� of the frame often give rise to laterally
directed rods towards their anterior that extend into the tergal area; these
could be displaced apodemes. Poole (1995) referred to basolateral apodemes of
the male eighth sternite as a trifine noctuid feature. Kitching & Rawlins
(1998) illustrated lateral rods in a much more posterior position in the
arctiid subfamily Ctenuchinae yet referred to them as continuations of the
antecosta of the sternite. Lödl (2000) illustrated the process of extrusion of
the sternal coremata in Eudocima and Phyllodes Boisduval.
The
tergite is usually considerably narrower than the sternite and is moderately to
strongly constricted just posterior to the apodemes. These characteristically
arise close together, from which point they are divergent, splayed. The tergite
broadens posteriorly from the apodemes to a rounded anterior margin that often
develops into a slight exvagination that would form a lobe or tumescence just
dorsal to the uncus. Even the narrow sclerotisation of the tergite can be
considerably reduced, often to just the apodemes and connected to a central
longitudinal band, e.g. in some Boletobiini.
The
sternite can also be modified further. It is considerably shortened, with the
lacuna vestigial, and developed into more of a W-shape in most Boletobiini and
many members of the Saroba group.
Within
the catocaline / ophiderine complex there are a few other types of modification
that do not fall within the �framed corematous� condition, as the apodemes of
the tergite are well separated and not splayed (see p. 189). Tribes and genera
with unusual modifications include the Hypocalini (p. 167), the Sypnini, and
the second sequence of miscellaneous genera from Ugia Walker to Hyperlopha Hampson starting
on p. 183.
Hyperlopha
and
Asta
Walker
in this second miscellaneous sequence contain some species that have
hair-pencils at the base of the abdomen, simpler than in the structure seen in
many trifine noctuid groups (Birch, 1972; Zilli & di Giulio, 1996) and not
necessarily homologous.
The
male genitalia
The
uncus frequently has an apical structure where a dorsal spur is superimposed on
a ventral bulbosity, referred to in the text following as the ball-and-claw
structure. The concavity between the spur and bulbosity is usually curved but
can be compressed into a narrow notch, e.g. in Lacera Guenée (Fig 132)
and Hulodes Guenée where it is used tentatively to define the
tribe Hulodini (p. 107). The more typical form is illustrated in Figs 81, 111,
150, 193 and 246. This form of uncus is encountered particularly but not
exclusively within the core Catocalinae.
Another
form of uncus that occurs in several of the genera in the �framed corematous�
sequence has the apex acute but not specifically of the ball-and-claw type. The
distinctive feature is a dorsal concavity, often extensive and associated with
flattening, that occurs just posterior to the apex. This concavity usually
contains a crest or tuft of hairs. This feature is seen in some of the Anobini,
in genera such as Plecoptera Guenée, Rhesala Walker and Tamba Walker, and in
some species of a few less diverse ones such as Gesonia Walker, Lopharthrum Hampson and Parolulis Hampson, the
last two being in the Saroba group with Tamba. In some genera the uncus has a
sharp peak or elongate dorsal process at the basal end of the concavity, e.g.
in Plecoptera, but some other
genera such as Chorsia Walker (p. 405) contain a few species with a similar
dorsal process. The structure in Rhesala can become
extremely elaborate, e.g. in more easterly species discussed and illustrated by
Holloway (1979).
A well
developed scaphium is also found in many of the core Catocalinae but is not
unique to them, being particularly massive in genera not included such as Asta Walker (p. 193)
and Pantura Moore (p. 195), and, displaced distally along the
ventral part of the uncus, is a diagnostic feature in the Scoliopterygini. At
its distal end the scaphium often partially encircles a pocket in the anal tube
that receives the apex of the uncus (e.g. Figs 247, 324, 682) or is apically
bifid at that point. The pocket in the anal tube is found in genera that lack a
strong scaphium, e.g. in Figs 518 and 528.
Fibiger
(2003) suggested that the length of the tegumen relative to the vinculum was a
useful feature for distinguishing core Catocalinae, these being characterised
often by the greater length of the vinculum. To some extent this is true, but
the character appears to be very plastic, with different states occurring
within genera such as Bastilla Swinhoe and Lacera Guenée. A long
vinculum occurs in the crameri Moore group of the former and the procellosa Butler group of
the latter, but in other species the tegumen is equal to or longer than the
vinculum. Amongst the Bornean core catocalines, a long vinculum is evident
particularly within the Catocalini, Ophiusini, Ercheiini and Pericymini, but
within the Ophiusini a longer tegumen occurs in genera in all three of the
lineages discussed on p. 40: Thyas Hübner, Achaea Hübner and Chalciope Hübner. A long
vinculum occurs in the Hypopyrini, but the tegumen is generally longer in the
Erebini. A long vinculum also occurs in genera of the �framed corematous�
sequence that are assigned by Fibiger (2003) to his Calpinae, e.g. Tamba (noted by
Fibiger) and Xanthanomis Hampson, and also Chrysopera Hampson.
Processes
from the tegumen also occur more frequently in the core catocalines, often
accompanied by bilateral asymmetry that can extend also to the valves and their
processes. Fibiger (2003) stated that all [European] species of Catocalinae
show some asymmetry of the male genitalia. This is seen particularly in genera
of the Ophiusini but also occurs in the Catephiini, Toxocampini and
Ommatophorini, and at least some species of genera such as Speiredonia, Ericeia Walker, Pandesma Guenée and Papuacola Hampson.
However, the feature is not restricted to the core catocalines but occurs more
rarely elsewhere, e.g. in genera such as Hamodes Guenée and Hyperlopha, and in the
�framed corematous� sequence, e.g. in some of the more robust taxa of the Saroba group.
The
articulation of tegumen and vinculum is almost always simple, without
interpolation of a pleurite or paratergal sclerite as described in more detail
on p. 25 as a feature for identifying genera better placed with the trifine
noctuid subfamilies. Interpretation of this character is difficult, and the
occurrence of a structure that could be a pleurite was noted in some genera or
species of the �framed corematous� sequence: genera of the fifth miscellaneous
sequence such as Cultripalpa Guenée and Pleurona Walker; members
of the Saroba group such as some species currently in Saroba Walker, Trichoblemma Hampson, Lopharthrum Hampson and Parolulis Hampson; and
genera of the sixth miscellaneous sequence such as Platyjionia Hampson, Leptocola Gen. n., Tamsia Roepke, Rhynchodina Hampson and Avittonia Hampson. Such a
structure was also noted in Varicosia Hampson of the first miscellaneous
sequence, within the core Catocalinae.
Another
feature noted in the region of the articulation of the tegumen with the
vinculum was a tendency of the former to loop interiorly into the diaphragma,
curling up dorsally. This feature was noted in the Boletobiini (e.g. Fig 794)
and some other genera such as Tamsia (Fig 820). Again
its nature and occurrence are difficult to interpret (e.g. in relation to the
transtilla structure described for the Catocalini by Fibiger (2003), discussed
at the end of this section; the groups just mentioned fall outside his
Catocalini), and so a full survey was not undertaken.
The
juxta may provide some features of phylogenetic significance, but these are
once more difficult to interpret with clarity. Many of the genera throughout
the text following have a juxta that is referred to in the text as being of the
inverted �V�,�Y� or �X� type (Holloway et al., 2001; Holloway
& Miller, 2003; Fibiger, 2003), all variations on a theme that, at its more
basic, consists of a sclerotised band arising on each side from an articulation
with the sacculus Fibiger (2003), and converging in the centre of the
diaphragma just ventral to the anellus. At this point the bands can become
fused into a central band (the �Y� shaped state) and then even diverge again around
each side of the anellus (the �X� shaped state) and sometimes become intimately
associated with it (e.g. in Catephia Ochsenheimer).
The
basic structure occurs very widely and can even be seen in genera currently in
the Hypeninae (e.g. Acidon, reviewed by Lödl (1998: fig 8)). However, as suggested by
Fibiger (2003), the more pronounced development of the feature such that its
vertical dimension is the longest tend to occur within the core Catocalinae,
but also, with modifications, in the Scoliopterygini outside the core
Catocalinae, and also in Ugia Walker and Heterospila Guenée . The
feature can be seen particularly clearly in Figs 12, 80, 102 and 131 for the
core Catocalinae and in Figs 294-298 for other groups.
Fibiger
(2003) indicated that the juxta in his concept of Calpinae was more usually
shield-like, heart-shaped, butterfly-shaped, ovate or generally a simple,
polygonal plate located more freely within the diaphragma. This is most clearly
seen in the Calpini as recognised here; in Eudocima the juxta is
bilobed, constricted centrally dorsoventrally, and usually with each side
produced into processes dorsally that can be very long and tapering, as
illustrated by Zilli & Hogenes (2002). If the basic �V� shaped juxta is as
generally distributed as suggested here, then these alternative forms of the
juxta may also be apomorphic (see also the Saroba group of genera
on p. 336).
The
valve structure, like the juxta, often shows variations on at least one theme,
but there are exceptions. The basic structure involves a series of processes
that traverse the valve from the base of the costa to the distal part of the
sacculus, often arising along an arch-like thickening. Fibiger (2003) suggested
that the costal process does in fact
represent a free costa, and certainly the dorsal margin of the more apical
portion of the valve rarely shows thickening when the more basal array of
processes is transverse and complex, as in the Ophiusini particularly amongst
the core Catocalinae; a detached costal process is also seen in Lacera but is otherwise
uncommon amongst the groups up to and including theAmphigoniini (p. 121).
However, if this is a feature of core catocalines as suggested by Fibiger
(2003), how should similar features in the �framed corematous� sequence be interpreted?
Hypospila
Guenée
and Tochara Moore have a prominent but relatively simple process
arising from the base of the costa, though with its base extending into the
valve. In the Saroba Walker complex of genera (p. 336), there is also an
array of processes running across the valve, though relatively more distally
than in the Ophiusini, particularly in the saccular area. The apex of the valve
is simple, tongue-like, often beyond a slight downward flexure at the junction
with the transverse processes. Conversely, in the Catocalini as recognised here
(p. 38) the costa is strongly sclerotised but still supports the membranous to
corematous distal part of the valve.
The
valve processes, therefore, may be more helpful in defining the Ophiusini
within the core Catocalinae and maybe other groups in the �framed corematous�
sequence, rather than in defining groupings at a higher level.
The
occurrence of prominent coremata on the valves may also be more applicable at
these lower levels. As in other families such as the Geometridae, their
occurrence is widespread but possibly relatively parsimonious. Amongst the more
strongly defined groupings recognised here, they are general to the Parallelia Hübner generic
complex in the Ophiusini (occurring also in Chalciope Hübner and Oxyodes Guenée of that
tribe), to the Erebini and to the Scoliopterygini, and the distal part of the
valve in Catocalini is somewhat corematous as described above. They are present
in Hulodes
Guenée
and Lacera of the Hulodini and in the Catephiini, tending to be
relatively distal in these. They extensive in Sympis Guenée. In the
�framed corematous� sequence they occur in some genera of the Episparis Walker group,
and also Claterna, Gracillina Hampson and Poliofoca Hampson. There
are small ones in Vestura Swinhoe and in some species of Rhesala.
The
aedeagus tends to be long and relatively slender, perhaps more so in the core
Catocalinae (Fibiger (2003)). The vesica is also generally more complex than in
other Noctuidae, with a number of slender diverticula of different sizes and
ornamentation, usually scobination or clusters and fields of small spines.
Massive cornuti are rare generally, though not uncommon in Calpini.
Not all
of the Catocalinae/Ophiderinae complex follow this pattern. The vesica is
relatively slender and simple in some Sypnini and most Scoliopterygini, simple
and somewhat recurved in Avatha Walker and some Tamba Walker, and
broader, more globular in various genera outside the core catocalines such as Heoeugorna Hampson, Phorica Walker, Pseudosphetta Hampson, Rhynchodina Hampson and some
in the Episparis Walker group.
Fibiger
(2003) suggested two further features of the male genitalia that he regarded as
contributing to diagnosis of a core concept of the Catocalinae. The first is a sclerotised
patch associated with the ductus ejaculatorius at its junction with the vesica.
Such a sclerite is not unique to the Noctuidae, though a globular feature in
this position is a synapomorphy uniting the Stictopterinae and Euteliinae
(Holloway, 1985: 165; Kitching & Rawlins, 1998). An analogous feature
occurs in the Lithosiinae (Holloway, 2001: 387). The second feature is
attributed to a more narrow concept of Catocalini equivalent to the groups that
are referred to the core Catocalinae concept here (i.e., in Europe, excluding
the Tytini and Armadini). It is an unusual transtilla that extends anteriorly
from the valve as a heavily sclerotised rod-like process that extends dorsad as
a �barely visible membranous string� that connects with its opposite number;
therefore the transtilla in the diaphragm is unsclerotised and difficult to
observe with certainty. It has not been surveyed in detail here.
The
female abdomen and genitalia
Fibiger
(2003) discussed several features of the female genitalia, particularly the
position of the ostium in relation to the seventh and eighth segments: core
catocalines typically have this in a relatively anterior position. He also
noted a degree of extensibility, with the terminal segments tending to be
retracted telescopically within the seventh segment at the apex of the abdomen.
This is a difficult characteristic to assess unless one has access to a
comprehensive sample of live or freshly killed material, as the drying process
may cause some of this retraction, particularly where the abdomen is broad and
tapers sharply. Fibiger (2003) noted in his Catocalini that the intersegmental
membranes were short ventrally and longer dorsally within the segments of the
genitalia, possibly correlated with the features discussed in the next
paragraph. A full survey was not undertaken for this work, given that much of
the examination of genitalia features was made on slide mounts prepared in the
past. Other features that can be associated with an extensile female ovipositor
in other groups, such as expanded intersegmental membranes, elongated apophyses
on the eighth segment or the ovipositor lobes, and modification of the
ovipositor lobes in a blade-like or acute, conical fashion, were not noted to
any extent in the majority of catocaline genera, though narrow, tapering
ovipositor lobes with a central strip of more marked sclerotisation were noted
in the Catocalini and a few individual genera as discussed on p. 38 and
elsewhere. Extreme extensibility is present only in Neogabara Wileman &
West (p. 450), which is well removed from the core Catocalinae.
The
more anterior positioning of the ostium is usually associated with modification
of the seventh segment, particularly reduction of the sternite posteriorly, but
the position of the former and degree of modification of the latter show at
least partial independence. Reduction of the seventh sternite involves
shortening from the posterior end and some tapering towards it such that it
becomes more triangular, though rounded at the posterior apex. Correspondingly,
the posterior corners of the tergite become pulled round ventrally and perhaps
somewhat produced laterally, though the whole abdomen becomes narrower also.
Thus the ostium can often be situated within the triple convergence of the posterior
corners of the tergite and the apex of the shortened sternite. The ostium
nevertheless varies in its position and the degree to which it shows fusion to
the ring of the eighth segment, or to the sternal apex of the seventh, or it
can be more or less free between the two; this is not always tightly correlated
with its actual position.
Core
catocaline groups usually have the ostium closely associated with the posterior
of a much reduced seventh sternite, the apex of which may be produced under it,
covering it, as a distinct plate that can be entire to deeply bilobed, and may
show asymmetry. This extension was termed the antevaginal plate by Holloway
& Miller (2003) and the lodix by Fibiger (2003). Though particularly well
developed in the Parallelia Hübner generic complex of the Ophiusini (Holloway
& Miller, 2003), it shows some degree of development in the groups treated
first in the systematic section up to and including the Hulodini (p. 117), and
in the genera from Erygia Guenée to Bamra Moore (pp. 130-149),
and may represent one of the most reliable features for recognition of a core
group of Catocalinae, as suggested by Fibiger (2003).
Fibiger
(2003) considered the condition of the base of the apophyses of the ovipositor
lobes to provide an autapomorphy for his Catocalinae. Generally, the base is
flat, diamond-shaped close to the ovipositor, but in the Catocalinae the
apophyses are rod-shaped throughout or only slightly broader at the base. This
character is hard to gauge, and a clear dichotomy between the core catocalines
and the remained of the genera covered here was not apparent to the author. The
lobes themselves may also present features that are definitive at a generic or
higher level: the narrowing and longitudinal reinforcement seen in Catocalini,
Catephiini and a number of other unrelated genera (p. 38); in Bamra Moore (p. 149);
in the Episparis Walker group of genera (p. 292).
In the
genitalia, some features of the ductus bursae, particularly the extent and
nature of the sclerotisation, e.g. in the Parallelia generic complex
(p. 53), may help define the core catocalines, the sclerotisation tending to be
more extensive and general in these (Fibiger, 2003). Fibiger also divides the
ductus bursae into the basal antrum and more distal ductus, these generally
both being sclerotised in core catocalines, with a short, transverse membranous
zone between them. This division is far from always apparent, and therefore the
whole is termed the ductus here. There are also problems (e.g. on p. 213 with reference
to the Scoliopterygini) in identifying where the ductus ends and the corpus
bursae begins, as the latter may have a neck of comparable width to the ductus.
The origin of the ductus seminalis from this joint structure may enable this
situation to be clarified but depends on the assumption that the ductus
seminalis is always at the base of the corpus bursae (see below).
The
shape and ornamentation of the bursa is variable, but a strong signum (or signa) is rare, and may often just represent concentration of a more general
scobination. Strong signa are seen particularly in the Scoliopterygini (p. 213)
Signa consisting of a scobinate patch are seen particularly in �Pantydia� metaphaea Hampson, Asta Walker, Bocula Guenée, Marcillada Walker, Xanthanomis Hampson, Poliofoca Hampson and Anticarsia Hübner, but can
be extended into longitudinal bands, e.g. in Hypocala Guenée, Hyperlopha Hampson, Taviodes Hampson, Focillistis Hampson, Psimada Walker and some Mecodina Guenée, Singara Walker and Tamsia Roepke. Bands of
spines or ribbons of scobination that ring the bursa, sometimes irregularly,
are seen in a relatively basal position in Buzara Walker, �Saroba� ceylonica Walker and
relatives, various Tamba species and Pseudosphetta. They occur centrally to
distally in Anoba rigida Swinhoe, Goniophila
excavata Swinhoe, the Mecodina diastriga Hampson group, Panilla and Platyjionia Hampson. In a
few genera in the sixth miscellaneous series, spining of the bursa is
extensive, the spines more of the size seen in many larentiine Geometridae.
Such spining is found in Aphypena Swinhoe, Rhesala Walker, �Loxioda� mediofascia Swinhoe, genera
related to Ilyrgis Walker, and Nolasena Walker. With the
exception of Buzara and possibly �Pantydia� metaphaea Hampson, all the
taxa mentioned above are outside the core catocalines.
The
point of junction of the ductus seminalis is usually at the base of the corpus
bursae, but exceptions include some Pangrapta Hübner, typical Hyposemansis Hampson, Panilla Moore, Rhynchodina, Egnasides Hampson, Semiothisops Hampson, Blasticorhinus Butler and Loxioda Warren species,
where it arises from the corpus bursae, and the Scoliopterygini where it is
often relatively more basal (p. 213).
Larval
features
Most
genera in the Catocalinae / Ophiderinae complex have larvae with the prolegs on
A3 and often A4 reduced or absent; they are semi-loopers, walking with a
looping motion but not as pronounced as in the Geometridae. A good sample of
genera was covered by Gardner (1947, 1948a) who placed them in his Group B,
with subdivision on the basis of the degree of proleg reduction. Such reduction
is not unique to the complex but is also seen in the Hypeninae amongst the
quadrifine subfamilies and in the Eustrotiinae / Acontiinae, Bagisarinae and
Plusiinae amongst the trifine subfamilies. Reduction of the A3 prolegs also
defines the Nolinae in the Nolidae (Holloway, 2003). Complete development of
the prolegs is seen in most of the Nolidae and trifine noctuid subfamilies, in
the Herminiinae and in the Stictopterinae + Euteliinae sister-pair. Reduction
is rare in other Noctuoidea but does occur in some Notodontidae (Holloway,
1983a: 5), e.g. the tribe Scranciini.
Its
more extreme occurrence in Geometridae has been shown by Young (pers. comm.; 2003) in a
molecular phylogeny to be somewhat plastic even within that family, with
occurrence of more complete proleg development represented in lineages
throughout the phylogeny she obtained. Therefore, as a guide to classification,
it needs to be noted with caution and in conjunction with other characters. The
degree of development of prolegs may be related to the life-style of the larva,
including its general build, the kind of vegetation on which it feeds and the
sort of crypsis it adopts when at rest.
An
example of this in the Bornean fauna may be presented by the Pandesmini (p.
101), where a firm grip of the substrate offered by a full set of prolegs,
together with robust, modified mandibles, may be essential adaptations for the
barkfeeding habit of the larvae.
Several
other instances of full proleg development occur in the traditional
Catocalinae/Ophiderinae. Some are noted on p. 26 and, in conjunction with other
characters of the adult and the presence in some of secondary setae in the
larva, are used to suggest placement with the trifine groups. Yet others are
listed by Gardner (1948a) in his Group C where reduction of the A3 prolegs, if
it occurs at all, is very slight. Those in the Bornean fauna are Anticarsia Hübner, Episparis, Fodina, Hypocala, Mecodina Guenée and the
Pandesmini (see above). In all of these genera except Hypocala the adult has a
scaleless lower clypeofrons, and most are allied to other genera or include
other species (Episparis) where proleg reduction occurs. Anticarsia larvae retain a
strong looping motion (p. 435). The case of Hypocala is discussed
more on p. 167. The anterior prolegs are not very strongly reduced in Catocala Schrank but the
first pair is not always fully functional (Bretherton et al., 1983).
Beck
(1992, 1996) advocated that much more attention should be paid to the
characteristics of early stages when establishing higher classification for the
Lepidoptera, particularly the Noctuidae. Gardner (1946a, b, 1947, 1948a)
provided details of chaetotaxy that would contribute to this. However, for the
tropical fauna in general, this is a counsel of perfection as our sample of
genera where details of early stages have been published is woefully small, and
those with even the detail offered by Gardner are much rarer. The manuscripts
of T.R.D. Bell contain a wealth of detail but were written before the major
studies of chaetotaxy were published in the middle of the last century. In this
series it has only been possible to present very brief and incomplete summaries
of this detail. Considerable rearing of noctuid larvae in the tropics,
including the Oriental, has been undertaken in the past few decades by J.E.
Rawlins and colleagues, but publication on this material is still awaited. A
flavour of what this may contribute is offered by the focus on early stages by
Kitching & Rawlins (1998).
Bell
had a concept of an �ophiusine� type of larva that is referred to frequently in
his descriptions, but it is hard to locate a clear definition of this. The
impression gained by the author is of a rather elongate larva with a slight
taper at each end and reduction of the two anterior pairs of prolegs. A
particular feature is presence of a pair of dorsolateral tubercles on A8, often
on a transverse tumidity or ridge. This is noted in Catocalini, many Ophiusini,
the Ercheiini, Catephiini, Hulodini and Amphigoniini, the Serrodes group and the
genera Ischyja Hübner and Bamra Moore. Most of
these fall within the core Catocalinae discussed on p. 38, but such tubercles
are also seen in the Erebini which do not, and are included by Fibiger (2003)
in his Calpinae. A transverse tumidity on A8 without definite tubercles can
also occur, e.g. in the Calpini, Hypocala and Hyperlopha Hampson.
Another
feature that was noted in several of Bell�s descriptions and which might repay
a deeper study is the presence of black or dark patches on the ventral surface
between the prolegs and often also between the true legs. These occur in
Ophiusini such as Thyas and Artena, Ercheiini, Catephiini, Erebini, the Serrodes Guenée group, Lacera Guenée, Spirama Guenée, Erygia Guenée and to
some extent in Ischyja Hübner and Hypospila Guenée. The
feature has also been recorded in Thalatta Walker in the
fourth miscellaneous series of genera.
The
value of larval and general biological features in higher classification of the
group has already been mentioned for the Pandesmini, where the adults of the
two genera involved bear little resemblance to each other (p. 101). This is
also evident in the Tinoliini (p. 122) where adults in Tinolius Walker appear to
be aposematic and those in Calesia Guenée mostly cryptic. The larvae,
however, share striking features such as an aposematic pattern with yellow
rings on black, loss of prolegs on A4 as well as A3 and elongation and
modification of the primary setae, though this is not pronounced in Calesia. The
host-plants of both genera are all in the Acanthaceae. There is also a striking
synapomorphy in the pupae as discussed in the next section.
Beck
(1996) presented a classification of European noctuids that was particularly
influenced by larval features and departs significantly from that of Fibiger
(2003) and the system adopted here. It has a concept of Catocalinae much
expanded from the core groups to include elements of the �framed corematous�
sequence such as the Boletobiini and the Pangraptini. The Scoliopterygini and
Calpini of the treatment here were placed as separate subfamilies,
Scoliopteryginae and Ophiderinae. The Aediinae and Bagisarinae were kept
distinct (see also p. 26).
Pupal
features
For
reasons given above for larvae, information on pupal characteristics is
fragmentary. Again, Bell gave very detailed accounts that are not summarised
here, though details of the strategy for pupation are; they contribute to
evidence for a relationship between the Calpini and Phyllodini (p. 261).
Gardner also provided an account of pupal features. He noted that the cremaster
often had eight hooked setae, but this appears to be general to many
Macrolepidoptera as discussed in the parts of this series covering the
Geometroidea. Gardner (1948b) noted that the trifine subfamilies, including the
�quadrifine� Plusiinae, Bagisarinae and Acontiinae [Erastriinae], have pupae
with the mesothoracic legs not extending to the eyepiece, whereas they do
extend to the eyepiece in other noctuid groups. There are, however, exceptions
amongst the latter, noted in the Anomis group, Oraesia Guenée, Arsacia Walker and Throana Walker.
One
feature that Gardner and many other authors have noted (e.g. Mosher, 1916;
Forbes, 1954; Bretherton et al., 1983; Kitching, 1984; Kitching &
Rawlins, 1998; Bell, MS) is the presence of a powdery or waxy, white, grey or
pale bluish bloom on the pupae of many Catocalinae (in the broad sense) that is
not soluble in alcohol. J.E. Rawlins (pers. comm.) urges caution
in using this as a definitive character for distinguishing a core Catocalinae
group before more is known of the chemical composition, fine structure, mode of
formation, relation to pupal surface ultrastructure and problems with assessing
presence as distinct from obvious abundance. Its presence as an obvious bloom
can vary within tribes and even genera, e.g. within the Ophiusini and between Avatha Walker (presence
noted) and Serrodes Guenée (absence noted). Some instances of recording
in the literature may reflect development of powdery mould rather than anything
inherent in the pupa and its formation, and some of the records of it listed
here based on rather tenuous observations, e.g. in Ericeia Walker and Homodes Guenée, may be
of this nature.
The
bloom has been observed in the Catocalini, most genera of Ophiusini, the
Pericymini (though not noted by Bell for two Indian species), Catephiini (the
type species of Catephia; Bretherton et al., 1983), Avatha in the Serrodes group, Lacera and possibly Ericeia in the tentative
concept here of the Hulodini, in Ischyja, Polydesma and Erygia, but not in the
Sypna
group,
the Ercheiini, Erebini, Hypocalini, Toxocampini, Amphigoniini, Calpini,
Phyllodini and Scoliopterygini except for Dinumma Walker where it
occurs.
It is
very rare amongst the �framed corematous� sequence of genera, though been
recorded in the possibly related genera Delgamma Moore, Hamodes and Hypospila, with possible
further records in the rather distinctive genus Homodes (p. 272) and in Attonda Swinhoe of the Saroba group (p. 336).
It also occurs in the Herminiinae according to Kitching & Rawlins (1998),
though it is not mentioned for the British herminiine fauna by Bretherton et al. (1983).
An
entirely different pupal feature, a dense golden pilosity, unites the genera in
the Tinoliini that are mentioned in the section above on larvae and described
in more detail on p. 122. This raises a question about the suggestion by
Speidel & Naumann (1995a) that the Tinoliini and the Palaearctic Euclidiini
(the subtribes Ectypina and Synedina of Fibiger (2003)) form a monophyletic
group, sharing a long, spiralled ductus receptaculi in the female genitalia
(usually removed in a normal genitalia preparation). However, the type genus of
the Euclidiini and a close Palaearctic relative both have non-pilose pupae with
a blue-grey bloom (Bretherton et al., 1983).
Misplaced
genera
�Quadrifine� genera probably better associated with the
�trifine� Noctuidae
Several
features are used here to identify genera historically associated with the
Catocaline / Ophiderinae and occurring in Borneo that may be better placed
within the trifine complex of subfamilies. The polarity of these character
states is often ambiguous, and some show potential homoplasy, so occurrence of
more than one of these features in any one genus can increase confidence in its
placement. The genera so identified will be treated in Part 13 of this series.
Poole
(1995) and Kitching & Rawlins (1998), as mentioned on p. 9, suggested that
a scaleless lower clypeofrons provided a reliable means of distinguishing most
quadrifine subfamilies from the trifine complex; it is only weakly evident in
Aganainae but is present in the Catocalinae complex, the Herminiinae,
Hypeninae, Stictopterinae + Euteliinae sister-pair, and a few smaller
subfamilies. Therefore the presence of extensive scaling on the clypeofrons
immediately above the base of the tongue is taken to indicate trifine
affinities in conjunction with other features. The Hypocalini (p. 167) have a
scaled clypeofrons, and the larvae have all prolegs fully developed, but
exclusion is not clearly supported by other features, so they are retained for
the time being within the Catocalinae.
Enlargement
of the counter-tympanal cavities in the metathorax was suggested by Speidel et al. (1996a) to be a
feature found widely in the trifine subfamilies. These cavities are moderate in
Bagisarinae and extraordinarily large in Aediinae and Tytinae (see below and
Yela & Kitching (1999)) as in higher trifines such as the Noctuinae and
Hadeninae. Only the Pantheinae resemble the Catocalinae, other traditional
quadrifine groups and the Nolidae in having them small. See also comments by
Poole (1995) on �Pocket IV� of the internal tympanic structure.
Speidel
& Naumann (1995b) considered the condition of the dorsal phragma of the
second abdominal segment to provide guidance on placement of noctuid genera;
such thoraco-abdominal features have been studied more generally in Ditrysia by
F�nger (1999). The plesiomorphic condition is probably the presence of a pair
of prominent and broad lobes extending from the anterior margin of the second
tergite (Figs 371and 388); such a feature is also seen in the Geometroidea
(though usually the lobes are smaller and narrower) and was noted particularly
in the Larentiinae by Holloway (1997: 120, 126) who termed the lobes apodemes.
The lobes are conspicuously present in the Catocalinae complex but strongly
reduced or absent in the trifine complex, possibly including the Pantheinae (Speidel
& Naumann, 1995b). This feature is therefore also useful for identifying
potential trifines amongst the Catocalinae complex.
Speidel
& Naumann (1995b), Speidel et al. (1996a) and
Kitching & Rawlins (1998) also reviewed the earlier work of Tikhomirov on
the musculature of the male genitalia, particularly the attachment of the valve
flexor muscle, M.4, at the articulation of the tegumen and vinculum. A
relatively more ventral attachment on the vinculum was noted by these authors
as an apomorphy for the Nolidae (Holloway, 2003). Poole (1995) and Kitching
& Rawlins (1998) discussed the occurrence of a pleurite or paratergal
sclerite that extends distally from the vinculum on each side and forms the
articulation with the tegumen; muscle M.4 attaches to this when it occurs. This
sclerite is secondarily fused to the tegumen in most Noctuoidea, including most
groups of the Catocalinae complex (Kitching & Rawlins, 1998; see also p.
17), and this could therefore be considered the plesiomorphic condition. Consequently,
its occurrence extensively in the trifine complex could be interpreted as an
apomorphic reversal and used to support identification of trifine genera within
the Catocalinae. Holloway (2001: 282, 386) also noted it as a potentially
useful feature for the higher classification of the Lithosiinae.
Other
features that, in combination with some of the above, can be indicative of
membership of the trifine complex (e.g. Poole (1995)) are, in the male
genitalia, presence of: a peniculus (a lobe bearing hair-setae) on each arm of
the tegumen, usually relatively ventrally; a corona of setae around the distal
margin of the valve; a relatively basal and simple harpe from the valve sacculus.
Speidel et
al. (1996a) defined a major clade of the trifine complex (but
excluding Bagisarinae and Pantheinae and several other subfamilies now
considered trifine) on the basis of possession of a structure they termed the
epaulette, situated between the conjunctiva and tympanal membrane on the
thorax. They noted the presence of an epaulette in the Aediinae and Tytinae.
Both these groups are regarded as true catocalines by Fibiger (2003), and the
latter group exhibits ambiguous features such as possession of a black terminal
patch on the hindwing between M3 and CuA1 and an antevaginal plate (lodix) in
the female genitalia, both regarded as catocaline characteristics. The Tytinae
do have a pleurite, though this was considered by Fibiger (2003) not to be
homologous with that of the trifines and to be the plesiomorphic condition rather
than apomorphic, as discussed above. The Aediinae, however, though sharing the
black terminal patch considered typical of the true Catocalinae by Fibiger
(2003), have an atypical juxta and several further trifine features as
discussed below. The epaulette is thought to be an adaptation for exclusion of
parasitic mites that specialize in feeding in the tympanal cavity of noctuids
(Treat, 1956).
Speidel
et
al. (1996a) noted a similar feature that they termed a
�pseudepaulette� in the tympana of Oxyodes Guenée (p. 78)
and Catephia Ochsenheimer (p. 84), but they considered this not
to be homologous with the feature seen in trifine noctuid subfamilies.
Occurrence
of four fully developed pairs of prolegs in the larva may also support a
trifine placement of a genus, though reduction of these is a highly plastic
feature that occurs widely outside the Catocaline complex (e.g. in Plusiinae,
Hypeninae, Nolidae and Geometridae) and can show reversal within it (p. 22). It
may be determined at least partially by the lifestyle of the larva. Possession
of secondary setae by the larva may also be an indication of affinities with
some of the more basal trifine groups such as the Acronictinae and Pantheinae.
Consideration
of all these features indicates that the groups discussed in the remainder of
this section may best be associated with the trifine noctuids.
The
Bagisarinae have been found to occur within the Oriental tropics as reviewed by
Holloway (1998, 2003) and Kitching & Rawlins (1998). Xanthodes Guenée will be
treated in Part 13; Chasmina Walker and Dyrzela Walker were
treated as Amphyrinae in Part 12 (Holloway, 1989) but are Bagisarinae. Two new
species of Calymniops Hampson have been found in Borneo. The type species,
trapezata
Moore
(Sri Lanka), and the new species show features that indicate close affinity to Dyrzela, particularly
aspects of the forewing facies and features of the eighth segment of the male
abdomen. It should be noted, however, that Poole (1995) considered that the
Bagisarinae were best associated with the quadrifine subfamilies.
Another
genus that may be best placed in the Bagisarinae is Ramadasa Moore. This has
a sparsely scaled clypeofrons; this scaling occurs immediately dorsal to the
base of the tongue, but is easily rubbed off. There is a pair of shallow but
broad lobes at the anterior margin of the second tergite. The male genitalia
share features with some chloephorine nolids as noted by Kobes (1997) and
discussed by Holloway (2003: 229), but also show parallels with those of Chasmina and Dyrzela. The male
eighth abdominal segment is complex but does not have the features discussed on
p. 14. The forewing shape, facies and finely textured, rather satiny scaling
are also comparable with those of some Bagisarinae, though the scales in the
leaden grey areas of the forewings are unusually two-toned in black and grey.
The
Aediinae have been mentioned above. They include the type genus, Aedia Hübner, many
species currently combined with Catephia Ochsenheimer,
and a number of very closely related or possibly synonymous Oriental genera
such as Eucatephia Hampson, Idicara Walker, Premusia Walker and Zarima Moore. All share
a rather distinctive radula-like structure dorsal to the anellus in the
structurally rather uniform male genitalia. Mosara Walker, included
in Callopistria
Hübner
by Holloway (1989), may also be related, as it shares this radula-like
structure (Holloway, 1989: fig 263), and association with Callopistria itself
(Eriopinae) should be investigated. The vinculum is much longer than the
tegumen in Aedia but only slightly so in Mosara. In the female,
the ostium is just posterior to the seventh segment but connected to the
eighth. The seventh sternite is only slightly reduced. Holloway (1976)
described two new species, susanae Holloway and pauli Holloway, in Catephia, associating
them with C. longiquua Swinhoe (Burma), now placed in Ecpatia Turner by Poole
(1989). Both are misplaced, and should also be combined with Ecpatia (type species dulcistriga Walker, Africa) combs. n., as they share
a very characteristic structure of the valve of the male genitalia, including
both a corona of setae and a basal harpe, and a prominent peniculus on the
tegumen. As in Aedia, the vinculum is longer than the tegumen. A few more species
listed under Catephia by Poole (1989) are probably better placed in Ecpatia, such as cryptodisca Hampson (W.
Africa), discophora Hampson (S. Africa), philippinensis Wileman &
West (Philippines) and sciachroa Hampson (Sri Lanka); melas Bethune-Baker
(New Guinea, Australia) was placed in Ecpatia by Edwards in
Nielsen et al. (1996). Several more species, as yet undescribed, have been
recorded from Borneo. The Aedia group and Ecpatia lack phragma
lobes on the second abdominal tergite. The larva in Aedia has all prolegs
fully developed.
Swinhoe
(1900) used a concept of Catephiinae that, whilst recognising Catephia in the broad
sense with the Aediinae included, also embraced the Arcte group and Cyclodes Guenée that are
discussed in the paragraphs below.
The
remaining genera probably all fall within the rather vague concept of the
Pantheinae as discussed by Holloway et al. (2001) as the
Pantheidae, following Kitching & Rawlins (1998). However it is probably
preferable to follow the results of the molecular analysis by Mitchell et al. (2000) and the
morphological study of Speidel et al. (1996a) and
treat the Pantheinae as a basal group within the trifine noctuid complex that
is probably paraphyletic. Poole (1995) grouped the Pantheinae among the trifine
subfamilies in a clade that also included the Acronictinae and the
Bryophilinae. All the genera discussed below lack phragma lobes on the second
abdominal tergite and have scaling on the lower clypeofrons. Many have a
relatively short third segment to the labial palps.
The
genera added to the pantheines by Kobes (1992) were stated by him to have setae
within the ommatidia of the compound eyes, though far less densely than in
traditional Pantheinae or Hadeninae. They also had dark rings on the tarsal
segments of the legs.
Kobes
(1992) associated the strikingly patterned genus Baorisa Moore, reviewed
by Behounek, Speidel & Th�ny (1996), with Trichosea Grote (the
Bornean representative was discussed by Holloway (1985)) in the tribe
Trichoseini in the Pantheinae. It has a large central corema in the male eighth
sternite as in Trichosea, and also shares with it an apical corona and basal
saccular harpe to the valve and a peniculus on the tegumen.
The
monobasic Sundwarda Swinhoe was suggested by G. Ronkay to the author to
be a pantheine. It lacks phragma lobes on the second tergite and has a scaled
lower clypeofrons. The eighth abdominal segment in the male is unmodified. The
male genitalia have the uncus short, robust, opposed by an extensive scaphium.
There is no paratergal sclerite. The valves are cleft into a saccular lobe and
a costal arm in a manner similar to several ennomine geometrid tribes such as
the Abraxini and the Macariini. The aedeagus vesica is generally tubular with
fine scobination. The legs are ringed but not prominently so.
Kobes
(1992) summarised the systematic history of the genus Trisula Moore,
historically (e.g. Nye, 1975) associated with the Ophiderinae, but currently
placed in the Pantheinae on the grounds of its reduced counter-tympanal hood,
anteriorly displaced basal position of vein M2 in the hindwing, and the fully
developed prolegs and presence of numerous secondary setae on verrucae in the
larva. This trifine complex placement is supported by the absence of phragma
lobes on the second tergite and by the presence of scales on the lower
clypeofrons.
The
male genitalia in Trisula have a typically trifine harpe on the sacculus but
the valves lack a corona. There is no paratergal sclerite. The uncus is
slender, strongly arched, with a scaphium. The aedeagus vesica is tubular with
a few small diverticula basally. The eighth abdominal segment is unmodified.
One species occurs in Borneo. The larvae have only been recorded from Ficus (Moraceae;
Robinson et al., 2001). Wiltshire (1990) referred the genus to the Trisulini,
but this appears to be the only usage of this family-group name and is not
accompanied by a valid diagnosis; it is therefore probably a nomen nudum (K�hne &
Speidel, 2004; Speidel & Naumann, 2005).
Arcte
Kollar
is one of the genera traditionally placed in the Catocalinae / Ophiderinae
complex, and has segregates in each group because some species have a spined
mid-tibia and others do not (e.g. Berio, 1959; Holloway et al. 1997, 2001). The
lower clypeofrons is scaled, the scales converging obliquely on the central
axis, and the phragma lobes on the second tergite are very shallow or absent.
The larva has all prolegs fully developed but lacks secondary setae. The facies
of the adults is striking, with cryptic, bark-like patterning on the forewing
and a somewhat iridescent, pale blue, sinuous fascia or two fasciae
postmedially on the otherwise dark hindwing.
In the
male, the antennae are filiform, smooth, and the eighth abdominal segment
appears to be a modification of the framed corematous type, with lateral rods
and a massive central corema on the sternite. The unusually smooth antennae and
the convergent scaling of the clypeofrons are also seen in the African genus Pseudoarcte Viette and the
Neotropical Pararcte Hampson; these also have similar forewing facies to Arcte. The group,
mostly the Arcte phylum of Berio (1959), would merit further study. Wiltshire
(1990) placed Arcte in the family-group Arcteini but this, as with Trisulini above,
would appear to be a nomen nudum (K�hne & Speidel, 2004; Speidel & Naumann,
2005). Arcte larvae may be specialist on Urticaceae and Ulmaceae (Robinson et al., 2001).
The
remaining genera all have a narrow, sinuous, pale fascia within the darker
border of the hindwing, though it is unclear whether this is homologous; in Cyclodes Guenée it curves
distad near the dorsum rather than basad. All lack phragma lobes on the second
tergite and have scaling on the lower clypeofrons. Kobes (1992) included Donda Moore in his
concept of the pantheines but excluded Belciana Walker on grounds
of differences of various external features and of the male genitalia, though
these are relatively diverse in form within the species he associated with Donda, and both
groups show a similar but not exclusive tendency for Malvales in their larval
hosts. Both have a male eighth segment of the framed corematous type, but this
is highly modified in Belciana. Both Donda and Belciana were considered
to be Acronictinae by Speidel & Kononenko (1998) and stated by those
authors to be �under revision by Sugi and Kononenko�; Yoshimoto (1998) also
placed Belciana in the Acronictinae.
Cyclodes
has
the lower clypeofrons sparsely scaled, the scales convergent at a central point
somewhat as in Arcte, and the male antennae are similarly smooth. The male eighth
segment could represent a modification of the framed corematous type, but the
sternite has only an ovate lacuna anteriorly rather than the strong corema of Arcte. The male
genitalia have what may represent both a peniculus and a paratergal sclerite,
and the valve sacculus terminates in a small, conical harpe. The larva has all
prolegs developed, but only primary setae are present, these set on robust
conical chalazae that ring each of the strongly annular segments (Kumar &
Naidu, 1992; H. Steiner, unpublished photograph). The larva feeds on the
inflorescences of palms (locs cit.; Robinson et al., 2001).
Ortopla
Walker
species often show strong sexual dimorphism, with males of the Bornean type
species, iarbasalis Walker, having a bite-like excavation in the apex of
the forewing. The hindwing illustrates particularly strongly the sinuous pale
fascia referred to above. The male eighth segment is a robust version of the
framed corematous type, the sternite with lateral rods. The male genitalia are
generally similar to those of Trisula except the valve
lacks a harpe.
Genera
and groups probably better associated with Herminiinae and Hypeninae.
Distinction
of the members of the �framed corematous� sequence of genera, particularly
those in the second half of this work, from other quadrifine groups such as the
Hypeninae and Herminiinae, is, at best, imperfect. As indicated on p. 15, the
�framed corematous� condition occurs in both these subfamilies, but their outer
limits are poorly defined. Lödl (1993; pers. comm.) recognises a
core group of genera around Hypena which does not show the framed
corematous condition though Harita Moore does (Lödl, 2000), but other
genera traditionally assigned to Hypeninae are proving difficult to coagulate
around this. Similarly, a major group of Herminiinae can be recognised, as
discussed by Owada (1987) and reviewed by Kitching & Rawlins (1998) and
Holloway et al. (2001). The herminiines are characterised within the
Noctuidae by possession of a prespiracular counter-tympanal hood which
separates otherwise rather similar genera such as Mecistoptera Hampson (Lödl,
1997) from Hypeninae such as Acidon Hampson and Hiaspis Walker (Lödl, 1998);
all three of these genera have a typical framed corematous male eighth segment
(Lödl, 2000). Hiaspis had, until the work of Lödl, been placed as a
synonym of Oglasa Walker (p. 402).
Several
genera included in the sixth miscellaneous sequence in the account that follows
are flagged as potential Hypeninae or Herminiinae, such as Nolasena Walker (p. 451;
Lödl (2000) commented on the unusually extreme modification of the male eighth
segment) in relation to Catada Walker, and Ananepa Hampson, Gonoglasa Hampson and Hepatica Staudinger (pp.
452-455) in relation to Mecistoptera or the Acidon complex. Egnasides Hampson is
indicated to have herminiine features on p. 440, and another traditional
ophiderine genus represented in Borneo, Ochrotrigona Hampson, was transferred
from the Ophiderinae to the Herminiinae by Owada (1998) despite its
postspiracular counter-tympanal hood.
It is
interesting to note that, in unpublished notes on the Hypeninae and Herminiinae
in BMNH, Miss A.E. Prout had made a selection of genera from the Ophiderinae
that she considered to be of �doubtful subfamily�. Many of these are included
here in the sixth miscellaneous sequence: Ilyrgis Walker and
allies, Hepatica Staudinger, Marapana Moore, Nicevillea Hamson, Nolasena, Pseudogyrtona Bethune-Baker
and Tadaxa Nye. She included Aphypena Swinhoe and Lithosiopsis Hampson of the
sixth sequence within her list of Hypeninae. The Throana group and
representatives of the Saroba group were also included in the
�doubtful� list, as were Cultripalpa Guenée and Poliofoca Hampson of the
fifth sequence and Brontypena Holland, Corcobara Moore and Pilipectus Bethune-Baker of
the first sequence.
Another
group placed traditionally in the Ophiderinae (Nye, 1975) that has been
excluded from them in recent works is the Rivulinae. This is essentially the
genus Rivula Guenée. It was placed with the Hypeninae by Kitching
& Rawlins (1999), but separated from them by Beck (1996), Speidel et al. (1996a) and by
Yela & Kitching (1999). Speidel et al. (1996b)
described distinctive features of the proboscis. Fibiger (2003) excluded the
Rivulinae from his concepts of Catocalinae and Calpinae; the type species of Rivula is the European sericealis Scopoli.
Therefore Rivula will be treated together with the Hypeninae and
Herminiinae in this series. Sugi (1987) also indicated that the genus should be
excuded from the Catocalinae and Ophiderinae. The widespread Oriental Daona mansueta
mansueta mansueta Walker was described from Borneo and is the type species of its
genus. It appears to be close in external appearance to Rivula species and will
therefore be treated in conjunction with that genus.
The use
of the family-group name Rivulinae poses nomenclatural problems, as it is also
used in Pisces, based on Rivulus Poey. Usage in Lepidoptera has priority,
but usage in Pisces probably has much greater frequency. For further comment,
see Speidel & Naumann (2005); ceding the name to ichthyology would pose
difficulties that might require submission of proposals to the ICZN for use of
Rivulainae as suggested by Speidel & Naumann should the moth group prove to
be monobasic (Speidel et al. (1996b) included Zebeeba Kirby, and Daona Walker may also
be included as indicated above).
Within
the traditional Ophiderinae (Nye, 1975) is a sequence of, for the group, very
small brown species. These are allied to Micronoctua Fibiger
(Fibiger, 1997) and are currently under study by Michael Fibiger who (pers. comm.) is
investigating the extent of what is proving to be a very diverse group, and assessing
its placement in the higher classification; he currently excludes them from his
concepts of Catocalinae and Calpinae. Bornean material and that collected by
the author in Sulawesi and Seram is currently on loan to him, and it is hoped
that the results of his studies can be included in a future part of this
series.
The
taxa relevant to Borneo that may belong to this group of �micronoctuids� are
Anachrostis
siccana Walker, Anachrostis fulvicilia Hampson, Tolpia
conscitulana Walker, Mimeugoa diapera Hampson (transferred to Neachrostia Hampson by Poole
(1989)) and Trigonochrostia excisa Hampson. All these species are currently
known only from their Bornean holotypes. The holotype of Tolpia
conscitulana, the generic type species, is a female with a damaged abdomen; Tolpia is potentially a
senior synonym of Micronoctua!
Host-plant
specialisations
Data on
larval host plants for the Catocalinae / Ophiderinae complex are very uneven in
their distribution. A high proportion is derived from agricultural or forestry
literature. More rarely, a more widely directed effort, such as that by T.R.D.
Bell in southern India, or a sampling programme without an economic basis
directed at selected plant families, such as that in New Guinea by Novotn� et al. (2002) and Miller et al. (2003), can
provide a less biased sample or fill in gaps in the coverage of plant families,
as does haphazard, �hobby� collecting of larvae. Also, published host records
inevitably are given equal weight and it is difficult to winnow out plant misidentifications
from observations of genuine but rare occurrences, such as those for Rubiaceae
and Euphorbiaceae as compared to those for Leguminosae from the New Guinea
programme for Plecoptera violacea Pagenstecher noted on p. 282.
Quantitative data of this nature are also very rare for the region.
All
these factors should be borne in mind as background to the comments in the
following paragraphs, which are intended mainly to provide a summary guide to
the greater detail given in the main text; some of the host associations are
based on very limited data.
The
economic bias may be at least partly responsible for the very large number of
indications of preference for Leguminosae, as the family is important in both
agriculture and forestry, and its species are often dominant in natural
vegetation types (though this in itself can lead to a greater diversity of
herbivores). Within the core catocalines, the tribes Hulodini, Hypopyrini,
Pandesmini, Pericymini, Ommatophorini (one record) and Toxocampini, and the genus
Erygia
Guenée
are all predominantly recorded from the family, and several open habitat taxa
in the third section of the Ophiusini (genera Mocis Hübner and Trigonodes Guenée) are
closely associated with Leguminosae and Gramineae, a pairing also exploited by Anticarsia Hübner (p. 435).
Dinumma
Walker,
tentatively included in the Scoliopterygini (p. 213), goes against the trend in
that tribe for Malvales, being recorded only from Leguminosae.
In the
�framed corematous� sequence of genera, legume preference is shown by the
Anobini and the genera Arsacia Walker, Blasticorhinus Butler, Hamodes Guenée, Hypospila Guenée, Lithosiopsis Hampson and Rhesala Walker, with
single but unique records from the family for Oglasa Walker, Pseudosphetta Hampson, Raparna Moore and Rema Swinhoe. The
only record for Bocula Guenée (p. 199) is from the Leguminosae.
There
is some specialism at a tribal level in other families, for example the
Scoliopterygini on Malvales and the possibly related (p. 261) Calpini and
Phyllodini on Menispermaceae, a family containing many tropical vines. In each
of these cases some other plant families may be exploited, particularly in
temperate latitudes where, e.g. for Menispermaceae, potential hosts may be
rarer. In the tropics, the calpine genus Eudocima Billberg has
taxa that also exploit other families more rarely such as Berberidaceae and
Lardizabalaceae; at the Pacific end of its range, Eudocima
phalonia Linnaeus has adopted the legume Erythrina as an
alternative host from Menispermaceae. The genus Avitta Walker has also
been recorded only from Menispermaceae.
Specialism
at a tribal level also occurs on a few other plant families such as Tinoliini
on Acanthaceae, the Hypocalini predominantly on Ebenaceae and the Serrodes Guenée group mostly
on Sapindaceae, the exception being Anereuthina Hübner on
Palmae. The few records for the Sypnini are from Fagaceae or Rosaceae. Some
lineages of the Achaea / Parallelia section of the
Ophiusini feed predominantly on Euphorbiaceae (see also Holloway & Miller
(2003)), a family also utilised by Attonda Swinhoe. Several
genera of the Episparis Walker group, such as Egnasia Walker and Claterna Walker, the Throana Walker group
(e.g. Throana and Nagadeba Walker) and
within the Pangraptini (p. 325) and the fifth miscellaneous sequence, such as Pleurona Walker, show
preference for the Rubiaceae.
Potential
generic specialism, indicated by single or a small number of records in most
cases, is indicated for Sapindaceae by Oxyodes Guenée, Sympis Guenée and Tathodelta Hampson, for
Apocynaceae by Fodina Guenée, for Connaraceae by Delgamma Moore and for
Lauraceae by Asta Walker, Pantura Moore, Marcillada Walker and Pilipectus Bethune-Baker.
The
Boletobiini (p. 373) are exceptional within the Catocalinae/Ophideriinae
complex in feeding on the tissues of fungi (particularly bracket fungi) growing
on trees, rather than on green plants.
Fruit-piercing
and blood-sucking by adult moths
The
propensity for fruit-piercing and blood-sucking in adult moths of many core
catocalines and of the tribes Calpini, Phyllodini and Scoliopterygini has
already been mentioned on p. 10 where the modifications of the tongue
associated with these habits are described.
The
fruit-piercing habit has been studied particularly in south-east Asia by
Bänziger (1982, 1987), who divided moths into primary and secondary fruit
piercers according to their ability to pierce fruit skins of varying toughness
and thickness, secondary piercers often using the holes made by primary
piercers to gain access to the juices of tougher fruit. A third group imbibes
sap superficially from damaged fruit. The capacity to pierce is related to
proboscis modification, those moths with erectile barbs being the most
proficient, able to penetrate the toughest skins such as those of citrus fruit
and of litchi, longan and their relatives. These primary piercers are found in
the Calpini, Phyllodini, Scoliopterygini, the Ophiusa section of the
Ophiusini, in the Pericymini and in Serrodes Guenée and Ischyja Hübner. Some
members of the Achaea / Parallelia section of the
Ophiusini are primary piercers, though most are secondary ones. Most of these
primary piercers have been observed (Bänziger, 1982) to have an antiparallel
motion of the stylets that generates a ratchet effect among species with barbs
that assists penetration.
Bänziger
(1982) noted that species in Platyja Hübner and
allied genera that are primary piercers do not have barbs, but the tip of the
tongue is very sharp and the rest is strongly sclerotised. He also included Sarobacala
albopunctata Semper (p. 147) in this category.
Blood-sucking
has been observed (Bänziger, 1968, 1980, 1983, 1986, 1989a) mainly in the
calpine genus Calyptra Ochsenheimer in south-east Asia, where about half of
the 17 species recognised by Bänziger (1983) to belong to the genus have been
recorded as blood-suckers. The usual victims are large, slow-moving mammals
such as perissodactyls, artiodactyls and even elephants, though they will also
attack humans. The mechanism for blood-sucking was described in detail for one
species by Bänziger (1980).
The
genus Calyptra provides an overlap between the suite of
fruit-piercing groups referred to earlier and those from a much more diverse
group of moth families that drink from lachrymal secretions and fluids on
carrion (e.g. Bänziger, (1973), and reviewed by Holloway et al. (2001)).
Biogeography
and ecology
The
complex as a whole is pantropical and attenuates sharply in diversity with
latitude. The groupings recognised here vary in their representation across the
tropical regions and in their penetration of temperate latitudes. There are
many family-group names attached to New World genera, and a preliminary survey
of male and female genitalia of the type taxa for these names indicates that
many may be unique to the New World, but investigation of relationships of
these to Old World groups is at a rudimentary stage. The core Catocalinae are
well represented, as are the Scoliopterygini, Calpini and Pangraptini as
defined here. The Boletobiini also include a new World component. However, it
is unclear whether the tribes Erebini, Hypocalini, Sypnini and Anobini, and the
Episparis
and
Saroba
generic
groups have close New World counterparts. Penetration of temperate latitudes,
indicated by the European fauna as recognised by Fibiger (2003), is restricted
to the core catocalines, especially the Catocalini, Ophiusini, Catephiini,
Toxocampini, Pandesmini and Pericymini as recognised here, and limited
representation of the Scoliopterygini, Calpini and Boletobiini (essentially the
type species of the type genera of these groups).
The
matrices in Table 1 provide biogeographic versus ecological profiles of the
major groups recognised. It is not always clear for records for Thailand
whether species otherwise only recorded in Sundaland extend north of the
Isthmus of Kra; such species are recorded under the �Sundaland� category in the
matrices.The first four, the Ophiusini, Serrodes group, the
Scoliopterygini (see also p. 213); and the Calpini with the Phylodini,
relatively low endemism and a high proportion of more widespread often highly
dispersive and migration species and, relatively, those associated with
disturbed forest and open habitats. Many of the latter are pests in agriculture
or forestry, particularly, as discussed in the previous section, the piercing
of fruit. All show some degree of larval host specialisation. The profile for
the Hulodini is not illustrated but is similar.
The
Scoliopterygini have the highest proportion (about 45%) of extensively
widespread Indo-Australian tropical species. This can be seen particularly in
the genus Rusicada (p. 224) as illustrated by the preliminary
phylogenetic hypothesis in Fig 2 on p. 230, that includes two species that are
of this nature, two species-pairs and possibly one further clade of more
localised species. The recorded host-plants are predominantly from the
Malvales, some taxa of which are particularly characteristic of disturbed or
open habitats, e.g. the genus Hibiscus. In this there
are parallels with the subfamily Bagisarinae, discussed earlier on p. 26 and by Holloway (1998). Some
Indo-Australian bagisarine genera are similarly widespread such as the Xanthodes generic group
and Chasmina Walker (Holloway, 1989: 186). It is interesting that
a definitive feature of the Bagisarine, appendiculate crochets in the larva is
also seen, probably convergently in Cosmophila Boisduval in the
Scoliopterygini.
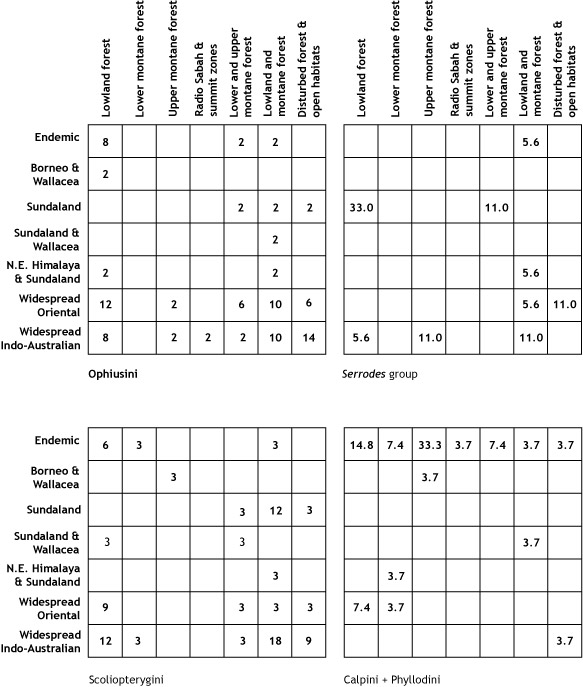
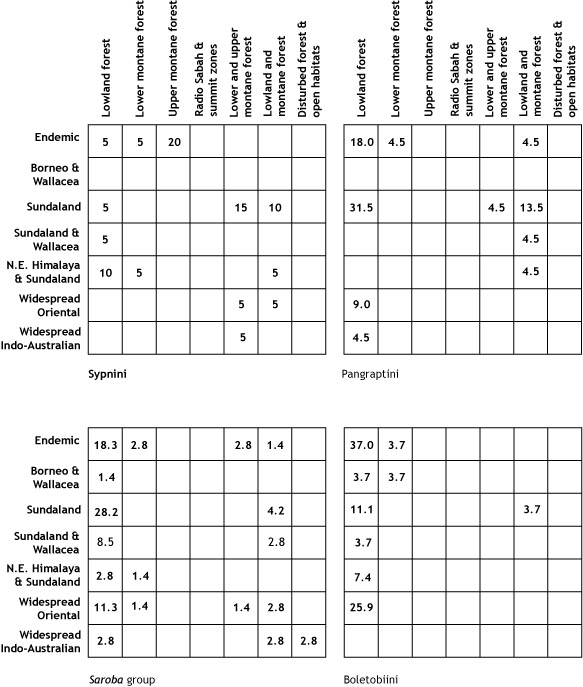
Table 1. Percentage of species for Catocalinae groups amongst
various biogeographic and ecological categories as discussed in the
text.
Apparently
widespread species should , however, be treated with caution, as a number have
proved on closer examination to be species complexes, very often being found to
break up into more localised allopatric taxa, particularly in the more eastern
extent of the range. Zilli & Hogenes (2004) illustrated this with a number
of east-west species-pairs, and cases requiring further investigation were
identified here for Macaldenia palumba Guenée (p. 68), Bocula sejuncta
Walker
(p. 200) and Hyperlopha cristifera Walker (p. 212).
Conversely,
named taxa distributed across the Indo-Australian tropics may prove to be
conspecific, exemplified here by Savara latimargo Walker (p. 220).
The
second four profiles in Table 1differ considerably from the first four. All
show much higher endemism, and also restriction of species to Sundaland. The
Sypnini are distinguished by high (over 50%) representation exclusively in
montane zones. The other three consist almost entirely (90% or more, though
many Pangraptini occur over a wide altitude range) of lowland species and
represent the predominant profile of groups in the Bornean fauna (Holloway
& Barlow, 1992; Holloway & Nielsen, 1998).
|