|
Erebus
ephesperis Hübner
Nyctipao
ephesperis Hübner, 1827, Verz. bekannter Schmett.:
272.
Phalaena
crepuscularis Cramer, [1777] 1779, Uitlandsche Kapellen,
2: pl. 159, fig. A, praeocc.
Nyctipao laetitia Butler,
1879, Illust.
typical Specimens lepid. Heterocera Colln
Br.
Mus.,
3: 26.
Erebus
malanga Swinhoe, 1918, Ann. Mag. nat. Hist. (9),
2: 85.
Erebus
niasana Swinhoe, 1918, Ann. Mag. nat. Hist. (9),
2: 85.
Erebus
ephesperis Hübner; Holloway, 1976: 29; Kobes, 1985: 24.
|

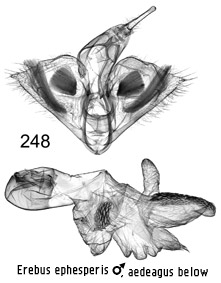
Erebus
ephesperis ♂
|
|
Diagnosis.
There is only limited sexual dimorphism, both sexes having a similarly
variegated brown pattern to the wings, though the males tend to be slightly more
strongly variegated, with a warm buff distal edging to the medial line. This
medial is strongly flexed round the ocellar mark and then passes obliquely
beyond it to form a band on the hindwing basal to the medial that itself links
with a buff band on the abdomen.
Taxonomic
note. The species is the most westerly member of a species group that
extends east to Vanuatu; this was reviewed by Fletcher (1957), who also divided ephesperis
into
four subspecies; the typical race occurs in Borneo. It can be distinguished from
other members of the group by its slender, flattened uncus and from crepuscularis, which flies with it in Java, by
having two clusters of spines in the aedeagus vesica rather than one.
Geographical
range. Oriental Region to Taiwan, Japan, Sundaland and Lesser Sundas
east to Timor.
Habitat
preference. This is essentially a lowland forest species like caprimulgus,
all recent records being from below 500m except one from 1200m at Bundu Tuhan on
the slopes of G. Kinabalu.
Biology.
The striking larva is illustrated by Mutuura et al. (1965)
and Sugi (1987) and also described (as crepuscularis)
by Gardner (1947) and Bell (MS). The prolegs on A3 are vestigial those on A4
reduced, but the rest are developed. A8 is humped dorsally. The head is
longitudinally banded with pale and medium rufous brown, but this banding fades
away into T1, and the dorsal surface posterior to this is a similar rufous brown
that is finely and variably blotched with darker brown in a rather reticulate
manner, the reticulations being greyish mauve laterally on A1 and A2 and
dorsally in bands across A7 and A8. The undersurface is dark brown that
interacts irregularly with the paler dorsal areas to give a disruptive effect:
strong over the thoracic segments, absent over A1 and A2, then recurring and
broadening dorsally over A3 and A4, the boundary then descending and becoming
sharp rather than diffuse over A5 and A6 (the dorsal area is much paler in this
section). There is a detached, oblique, irregular dark brown streak laterally
from A6 to A7, and the ventral part posterior to this is again dark brown. On A1
there is a conspicuous dorsolateral brown ocellus that is finely ringed with
mauve and black, and a centrally disrupted, transverse black bar within a paler
halo dorsally at the junction of A1 and A2. Bell (MS) noted that the larva bore
some resemblance to those in Eudocima
Billberg,
and stated that the segments were well defined and that the short primary setae
were set on chalazae.
Bell observed that the larva is sluggish and retiring, feeding by night, lying
by day on the stems of the host plant, near the roots, or on the ground or a
branch of the tree that supports the host vine. The defensive posture involves
bunching and curling the anterior part of the body as in Eudocima,
but the anal section is never lifted off the substrate. Pupation is in a cell
amongst leaves drawn together. The adult resting behaviour is typical of the
group (see E.
macrops Linnaeus
below).
The host-plant recorded (also Miyata, 1983) was always Smilax
(Smilacaceae).
<<Back
>>Forward <<Return to Content Page
|