|
Paectes
cristatrix Guenée
Ingura cristatrix Guenée, 1852, Hist. nat. Insectes,
Spec. gen. Lepid. 6: 313.
Paectes cristatrix Guenée;
Holloway, 1976: 17(part).
Diagnosis. This and the next species have strikingly marked forewings with a
pinkish tinge over the paler bands. P. psaliphora has the dark areas at
the dorsum more greenish and the posterior margin of the basal pale mark is
convex, directed at the thorax, rather than concave, directed at the dorsum as
in cristatrix. The male genitalia of the two species differ mainly in the
aedeagus vesica and its ornamentaion: in cristatrix there is a small
subbasal lobe, a slender central cornutus, and the vesica tapers slightly to a
larger, blade-like distal cornutus at the base of the ductus ejaculatorius; in psaliphora
the vesica is more globose with, in Bornean specimens, three cornuti as
illustrated (in typical New Guinea psaliphora the vesica has only two
cornuti). P. cristatrix can also be defined by the female genitalia where
there is a distinctive double angle to the distal limit of the zone of
scobination in the basal half of the bursa; in psaliphora this part of
the bursa is more globular, much less sclerotised or scobinate.
Taxonomic notes. Both cristatrix and psaliphora belong to a complex of
species that ranges through the Indo-Australian tropics to as far east as Samoa.
The geography of these species is interesting (Fig. 7) so they are discussed
here in some detail. Other members of the complex are:
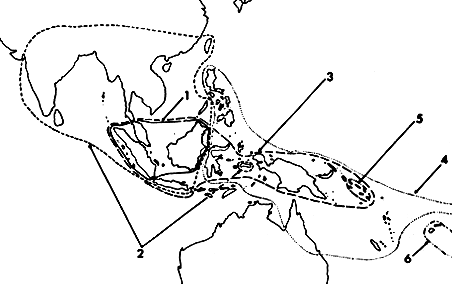
Figure 7. Distribution of species of the cristatrix group of Paectes. 1.
P. poliotis, 2. P. cristatrix, 3. P. psaliphora, 4. P.
cyanodes, 5. P. languida, 6. P. fijiensis.
P. cyanodes Turner. This species ranges typically from Queensland to New Guinea and
the Solomons. The male resembles that of cristatrix though is slightly
less pink; the female has the pale areas more heavily suffused with mauvish pink
rendering the wing uniform in tone except for the posterior edge of the basal
patch. Apart from this sexual dimorphism the species is defined by two cornuti
in the aedeagus - a long basal one and a short distal one, typically broad but
varying in other races, at the base of the ductus ejaculatorius. In the female
there is a distinctive tongue-like process from the basal part of the bursa
copulatrix. A series of races ranges from the Philippines to Samoa.
In the Philippines, Sulawesi and Saleyer there occurs a form that is
similar in facies but with the distal cornutus slender and half the length of
the more basal one rather than less than a quarter, and with the basal part of
the bursa less scobinate. In the Kei Is. is a form with an extra cornutus
near the subbasal one. In New Caledonia and Vanuatu there is a race
lacking sexual dimorphism, both sexes being as in typical males (illustrated by
Holloway (1979), but referred erroneously to cristatrix). In the Sta Cruz
Is. there is a race larger than that in Vanuatu, darker, and with a degree of
sexual dimorphism, and in Samoa flies ssp. canescens Tams stat. n., a
large race with rather uniform facies in both sexes but a degree of sexual
dimorphism also. The eastern end of the distribution of cyanodes is of
interest as it is yet another example of the pattern running from the Solomons,
New Caledonia and Vanuatu to Samoa that excludes Fiji (Robinson 1975; Holloway
1979, 1983b).
P. languida Warren. This species is known from only two specimens from the Solomons.
The male forewing is shades of fawn with a very broad, oblique, paler medial bar
extending up to the postmedial loop. The female is smaller, darker, a mauvish
grey. In the aedeagus vesica the cornutus at the base of the ductus
ejaculatorius
is vestigial but there is one long slender one centrally and one slightly
smaller one nearer the vestigial one. The female has an angled sclerotised
right-hand edge to the basal part of the bursa.
P. fijiensis Robinson stat. n. This species flies in Fiji and Tonga. It resembles cristatrix,
though somewhat larger, and was originally described as a subspecies thereof
(Robinson 1975). It is here given specific status because of its isolation from cristatrix
and relationship to other species in the south-west Pacific. The male
aedeagus vesica has a short cornutus at the ductus ejaculatorius as in cyanodes but opposite this there is a
single slender cornutus on one side and two long slender ones on the other. The
basal part of the bursa copulatrix is slightly asymmetrical and ringed by a
diffuse band of scobination at two thirds.
The species of the complex overlap each other considerably, suggestive
of extensive dispersal during the evolution of the group. Using the guidelines
of allopatric divergence and parsimonious dispersal suggested by Holloway (1982a), the evolution of the group may have proceeded as follows: (1) isolation
of cristatrix and poliotis ancestors in mainland Asia and
Sundaland respectively; (2) spread of the cristatrix ancestor to as far
east as Fiji; (3) isolation and divergence of cristatrix, psaliphora,
cyanodes, languida and fijiensis in mainland Asia, Sundaland,
somewhere between Sulawesi and New Guinea, the Solomons and Fiji respectively,
in allopatric but possibly contiguous ranges; (4) a second dispersal phase
leading to the modern ranges of the five species; (5) minor divergence again,
especially of cyanodes. If this narrative of events is correct then, as
with Aplotelia, dispersal has been almost entirely from west to east, and
more particularly involving the species associated with larger land areas
initially. Even if Melanesia is taken as the area of ancestral isolation of the cristatrix
group in (1), this only adds one easterly range extension.
Geographical range (cristatrix). Oriental tropics to Sundaland and Palawan;
also recorded from Ceram.
Habitat preference. The only specimen taken during recent surveys was at
1930m on G. Kinabalu.
<<Back
>>Forward <<Return
to Contents page
|