|
The Boarmiini are treated here in a very broad sense, including also the
following family group names: Amphidasini, Ascotini, Bistonini, Braccini,
Bupalini, Cheimatobiini, Cleorini, Eubyjini, Gnophini, Hyberniini, Melanolophini,
Milionini, Selidosemini, Sionini and Theriini.
The group shares with the Macariini, Eutoeini and Cassymini the derived
character of the bifid pupal cremaster, but may be paraphyletic in relation to
them particularly in view of the more plesiomorphic cremastral condition seen in
Acrodontis (See Acrodontis
Wehrli). All these groups share possession of a fovea in the
male forewing of some genera, these genera being in a majority in all except the
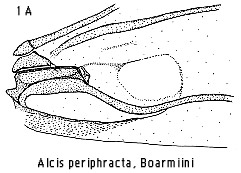
Cassymini. The structure of the fovea is distinct in each case (Fig 1). In the
Boarmiini it occurs as a depression, viewed from the underside, between the anal
vein and the fold of CuP. It is ovate to rounded-triangular, often with a
transverse fold marking the distal margin. The anal vein is often flexed
posteriorly to accommodate it, but in a more even, less sigmoid manner than in
the Macariini, resembling the Eutoeini in this.
 |
The male antennae are various, from fasciculate to strongly bipectinate
or quadripectinate. There is usually a transverse comb of setae on the third
abdominal sternite.
The majority of genera have rather cryptic, ligneous or lichenous wing
markings. Strong angling of the forewing postmedial distal to the cell is a
frequent feature (also in Macariini). But a number of genera, featured early in
the systematic arrangement following, are more colourful, more strikingly
patterned, and tend on the whole to have the forewing postmedial less
conspicuously angled. The former group coincides broadly with the Hypomecis Hübner
complex of genera discussed by Sato (1984a), and placed in the second half of
the arrangement.
The male and female genitalia provide only limited indications of
groupings within the tribe, and no unambiguous tribal features. In the male, the
valve is entire and usually has a marked cucullus at the apex of the costa,
strongly invested with setae, but this may be homologous with the dorsal portion
of the divided valve in the Eutoeini and Macariini. The sacculus is often
ornamented with spines or sclerotised processes bearing spine-like setae,
infrequent in the related tribes, and from this ornamentation a sclerotised
band, sometimes with further ornamentation, extends across the lamina of the
valve to the costa. Extreme development of this feature is seen in Bracca Hübner.
Uncus and gnathus are strong, the former often bifid. Coremata at the valve
bases or in the abdomen are not frequent. The aedeagus vesica has various
ornamentation, sometimes with very massive cornuti.
In the female, the signum is often of the typically ennomine dentate
mushroom form, but is modified in some genera or groups of genera: a transverse
flange in the Myrioblephara Warren group of genera; two longitudinal bars
of sclerotisation in Psilalcis Warren and allies; reduced, bicornute in Hypomecis,
Microcalicha Sato and a few other genera. The ovipositor and apodemes are
elongate, extensile in a number of possibly independent instances: in Hyposidra
Guenée and allies; in Biston Leach and related genera; in Cleora Curtis,
Ectropis Hübner and a few other genera in the Hypomecis group of
Sato, though a relationship to Biston is possible. All these genera
include a number of highly polyphagous species, though it is not clear how the
two features are correlated. The ovipositor is adapted for placing eggs in deep
crevices, e.g. in bark, in species where information is available.
The polyphagy just mentioned is often very wide, including both
angiosperms and conifers. Some other genera in the tribe show a high degree of
specialisation. These are noted in the Introduction (See
Introduction) and the section
following.
>>Forward
<<Return to Contents page
|