|
SUPERFAMILIES
DREPANOIDEA AND GEOMETROIDEA |
These two superfamilies as represented here include the majority of the
families that were united in the old concept of the Geometroidea (e.g. as
delimited by Fletcher (1979) and Holloway, Bradley & Carter (1987)). It was
broadly defined by the presence of abdominal tympanal organs, though excluded
the Pyraloidea that have them and included a number of small families that do
not. Minet (1983) made a detailed study of these tympanal organs and concluded
they had evolved several times independently and had distinct structure in each
lineage that was not homologous (see also Cook & Scoble (1992) and Scoble
(1992)). He divided the old Geometroidea up into several superfamilies that were
effectively monobasic, each containing a single family: Geometroidea,
Drepanoidea and Uranioidea. Subsequently (Minet, 1986, 1991), he has reunited
the Uranioidea with the Geometroidea and suggested that this grouping has a
sister-relationship with the Drepanoidea, indicated in particular by characters
of the pupal groundplan (Minet & Scoble, in press): strong anterior
extension of the forelegs; a transverse dorsal groove on A10. He has also
included some of the smaller families without tympanal organs: Epicopeiidae with
Drepanoidea, and Sematuridae (Neotropical, though including the southern African
Apoprogonidae) with Geometroidea. This arrangement is followed by Minet &
Scoble (in press) and in Fig 1.
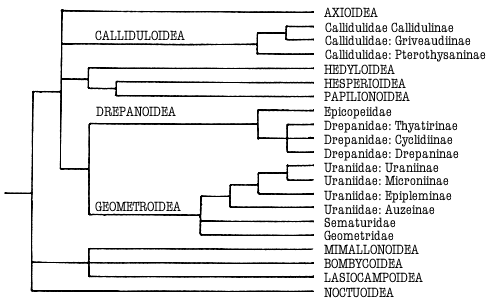
Figure 1. Phylogeny of the Macrolepidoptera as suggested by Minet
(1991) and Kristensen & Skalski (in press). Superfamilies are indicated in
capitals. Groups treated in this work are indicated in bold.
This new concept of the Geometroidea is defined (Minet, 1991) on: the
larval spinneret is shorter than the midline of the prementum; the ventral arm
of the tegula in the adult is more or less pointed and elongated. The latter
feature is also seen in some Epicopeiidae but deemed to have evolved
independently.
The Drepanoidea are defined by Minet on four apomorphies: a
characteristic of the larval mandible and its setae; at least one secondary seta
associated with L3 on segments A1-8 (though present in many geometroids except
Sematuridae); concealment of the femur of the pupal foreleg; adult abdomen with
prespiracular sclerite laterally, connecting the first tergite with the sternite,
and modified into tympanal organs in the Drepanidae.
This system of classification will be followed here. However, some
further comments on the Epicopeiidae are merited. This small Oriental tropical
and subtropical family is not known from Borneo but is represented by one
species in Sumatra, Epicopeia battaka Dohrn. Though it shares the
apomorphies noted above with the Drepanidae, it also has features in common with
the Uraniidae, such as an unusual branching system for the forewing venation (Scoble,
1992; Auzeinae). Minet & Scoble (in press) also define the family on the
following characters: the compound eyes of the adult are reduced in size;
absence of a forewing areole; the pupa has anteriorly broad galeae. Most genera
have a pair of hair-pencils basally within the male abdomen: such structures are
also seen in the Cyclidiinae of the Drepanidae (See
Cyclidia
Guenée) and some Oretini
(Watson, 1965a).
The larvae secrete granules or filaments of wax, a further possible
family autapomorphy (Scoble, 1992). The pupal cremaster has eight hooklets, the
apical pair stronger. Pupation occurs in a weak cocoon on the ground.
Yen, Mu & Jean (1995) have suggested that there are three major and
quite distinct lineages in the Epicopeiidae. They indicate that further
observations are needed to support the concept for the family of Minet (1991).
The ecological and biogeographic representation of Bornean Drepanidae and Uraniidae
Figure 2 presents summary tables of the biogeographic and ecological
profiles of Bornean drepanids and uraniids, following the conventions of
Holloway & Barlow (1992) and Holloway (1994, 1997).
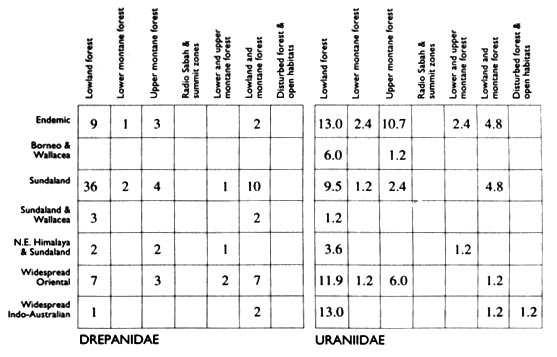
Figure 2. Percentage of species for Drepanidae and Uraniidae amongst various
biogeographic and ecological categories as discussed in the text.
In both groups species of lowland forests predominate, though the Uraniidae have
a slightly higher proportion of montane species (29% to 19%), and the Drepanidae
have many species ranging from the lowlands to montane zones (23%). The
Uraniidae also include more endemics and more geographically widespread species:
over half the Drepanidae are restricted to Sundaland but occur in areas
additional to Borneo.
Both families show preference for undisturbed forest, with larvae feeding on
trees and shrubs. Only one epiplemine, Dysaethria quadricaudata Walker,
has been taken in any frequency in disturbed forest or plantations (e.g. Chey,
1994; Chey, Holloway & Speight, 1997), and records of Drepanidae and the
uraniine Lyssa menoetius Hopffer in the lowland softwood plantation
survey by Chey were more frequent in secondary forest or the Eucalyptus
deglupta plantation where a secondary forest understorey was well developed.
In a comparison of four forest types in Peninsular Malaysia with various degrees
of disturbance or management, only uraniids were represented in any numbers, and
then in the forest types (associations D and E) that approximated most closely
to a natural state (Intachat, Holloway & Speight, 1997). In a comparison of
pairs of sites in undisturbed versus selectively logged forest types, Intachat
(1995) noted only a slight decrease in numbers of uraniid species in the logged
sites and a slight increase in drepanid species, so both groups can tolerate, or
maybe even benefit from, a moderate degree of disturbance. Neither family was
represented strongly in samples from areas of shifting cultivation in Seram,
though both were moderately diverse in samples for various undisturbed forest
types (Holloway, 1993).
With the mainly temperate and subtropical Thyatirinae excluded, the Bornean
drepanids represent 16% of the Oriental Region total given by Heppner (1991).
Similarly, Bornean Uraniidae represent 40% of the Oriental fauna.
|