|
Arctornis
Germar
Type species: v-nigrum Fabricius, Europe.
Synonyms: Cassidia Walker (type species obtusa
Walker) syn. n.; Chatracharta Walker (type species tortricoides
Walker) syn. n.; Ciaca Walker (type species urapterides Walker
= rutila Fabricius) syn. n.; Cobanilla Moore (type species marginata
Moore) syn. n.; Kanchia Moore (type species subvitrea
Walker, India) syn. n.; Lymantralex Collenette (type species heteroides
Collenette) syn. n.; Redoa Walker (type species submarginata Walker);
Scarpona Walker (type species ennomoides Walker = rutila) syn.
n.; Topomesa Walker (type species subinanis Walker = rutila)
syn. n.
This genus is extremely diverse in the Oriental tropics, particularly
Sundaland, and extends more weakly into the Palaearctic Region and to New Guinea
and Australia. Figures in Schintlmeister (1994) suggest that there are at least
46 species in the genus in Sumatra, and the figure for Borneo is approaching
twice this. Sketches of the genitalia of unidentified Sumatran species (Schintlmeister,
MS) sent to the author have enabled the ranges of some of the new Bornean ones
to be extended to Sumatra.
The genus includes as synonyms Redoa, following Heppner &
Inoue (1992) and Nielsen, Edwards & Rangsi (1996), and a number of smaller
Oriental genera such as Cassidia, Cobanilla, Kanchia, Lymantralex and Scarpona,
partly as suggested by Roepke (1948). There are differences in venation
across the genus, and there are also some more distinctly (and, in cases,
convergently) patterned taxa, but all have the same distinctive features of the
male genitalia: an unusual articulated arm, or harpe, often very long and
slender, arising from a pocket on the valve sacculus. There may be further
ornamentation to the valve margin. The aedeagus is usually short, often flanked
by a pair of distinctive lobes or digitate processes. The uncus is variable in
form, usually short, broad, sometimes bilobed. The male abdomen has tymbals.
The wings are usually pale-coloured, the majority of the species being
satiny white, sometimes with a greenish tinge, or fawn, yellow or shades of
brown. The satiny white species often have a rippled effect to the forewing,
generated by the distribution and orientation (e.g. ridging) of large, broad
scales relative to small, narrow, forked ones (see Holloway (1976)). Sometimes
the former are few or absent, giving the wings a translucent appearance. The
venation is variable, the forewing with or without an areole, and, when present,
variability in how it is formed: the key of Toxopeus (1948) relies particularly
on venation characters.
White and pale yellow species are often marked in various ways: rusty
brown on the forewing costa, labial palps and frons, and occasionally on the
fringes. There may also be a black discal spot, or spots on the legs. The wing
veins may also be darkened.
Association of males with females is very difficult, and it is
recommended that new taxa be based on males, as these tend to be much commoner
in light-trap samples. Some of the minutiae of markings, including those on the
palps and legs, may assist in this process, as may the length of the palps and
their segments. Therefore, the account of species following will focus almost
entirely on males. Females have genitalic features as in the tribal description
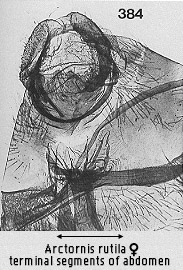
above, but show some variation in the ovipositor lobes. These can be simple,
rather square, or modified by shortening and slight scrolling into arcuate,
rather lip-like structures (e.g. Fig 384).
The diversity of the genus in Sundaland, particularly in undisturbed
lowland forest (Holloway, 1984), suggests it may, with the Nygmiini, be of
importance in monitoring the ecological health of such forests: some species
even feed on Dipterocarpaceae (see above and below), and further records of
association with this important family of rain forest trees might be predicted.
It is therefore unfortunate that much of the type material relevant to
Borneo for species described by Toxopeus (1948) cannot be located. The original
descriptions indicate it should be in the Muzium Zoologicum Bogoriense, but
several efforts (e.g. by H.S. Barlow and the author in conjunction with MZB
staff, as well as by Mackey (1984)) to find it have only been partially
successful (Mackey, 1984). Much Toxopeus material in other groups is still
extant, including manuscript holotypes in Nygmia and the Lasiocampidae,
but it is clear from his 1948 publication that the period during and immediately
following the 1939-45 war did not provide the stability conducive to taxonomic
research and the preservation of scientific material. Similar problems occurred
in Europe during the same period, e.g. in diatom taxonomy (Mann, 1998). The lack
of access to type material or its loss can provide a stumbling block to
nomenclatural stability in the group concerned, and any attempt to get round
this is likely to prove only partly satisfactory.
Toxopeus (1948) provided a key to his new taxa, obviously examined the
male genitalia of many of them, but did not illustrate these. The occurrence of
approximately twice as many species in Borneo as he was aware of means that the
characteristics given in the key may not enable one to identify reliably the
species for which type material is missing unless the features concerned are
highly distinctive, such as in species like arbor-christi and isabella.
Other Toxopeus species based on Bornean material where the type is lacking
are adusta, clavigera, erasmia, flaminea, linteola, salebrosa and satinata.
A few of these are represented by specimens in BMNH that have been labelled
by C.L. Collenette as having been compared with Toxopeus type material. These
will therefore be used to establish an identity for the names concerned: adusta,
clavigera and linteola. It is likely that some of the four remaining
species are redescribed in the species accounts following, but it is important
that studies of the genus in the region should not be impeded by a lack of
workable taxonomy, even if this should need nomenclatural adjustment later if
further types come to light. Some diagnostic features of the four Toxopeus
�unknowns� are given at the end of the species descriptions below.
The more distinctive species are treated first, followed by the main
mass of satiny white ones, where more detailed descriptions of head and leg
features will be included.
The larva of the type species is illustrated in Carter & Hargreaves
(1986). It has typically lymantriid secondary setae on verrucae, but those at
the posterior, directed backwards, and on the thorax, directed forwards, are distinctly
longer. This feature is also seen in the species illustrated by Wang (1993).
Possible tribal features have already been mentioned.
Because of the problems of establishing the identity of members of the
white group of species, host records associated with named species in the
literature should be treated with caution. The genus as a whole in its tropical
range has been recorded (Gardner, 1938; Sevastopulo, 1940, 1941; Kuroko &
Lewvanich, 1993; Wang, 1993; T.R.D. Bell, MS; unpublished IIE and FRIM records)
from the following hosts: Anacardium, Mangifera (Anacardiaceae); Durio
(Bombacaceae); Terminalia (Combretaceae); Balanocarpus, Hopea,
Neobalanocarpus, Shorea (Dipterocarpaceae); Diospyros (Ebenaceae); Elaeocarpus
(Elaeocarpaceae); Alseodaphne, Cinnamomum, Litsea, Phoebe (Lauraceae);
Lagerstroemia (Lythraceae); Medinilla (Melastomataceae); Nephelium
(Sapindaceae); Camellia, Gordonia (Theaceae).
Bell (MS) indicated that parasitism by tachinid flies was often heavy in
S. India.
<<Back
>>Forward <<Return
to Contents page
|