|
Eudocima
[Othreis]
phalonia
Linnaeus
[fullonia
Clerck]
Phalaena
Noctua phalonia Linnaeus, 1763, Centuria Insectorum rariorum:
28 (revived senior synonym of fullonia; see below).
Phalaena
fullonia Clerck, [1764] 1759, Icones Insect. rariorum,
pl. 48, ff. 1-4.
Phalaena
(Attacus) fullonica Linnaeus, 1767, Syst. Nat. (ed.
12): 812.
Noctua
dioscoreae Fabricius, 1775, Syst. Ent.:
593.
Phalaena
Noctua pomona Cramer,
[1775] 1779, Uitlandsche Kapellen, 1: 122.
Ophideres
princeps Boisduval, 1832, in d�Urville Voyage
de D�couvertes de l�Astrolabe, Pt 1 (Lep.): 245.
Ophideres
obliterans Walker,
1857, List
Specimens lepid. Insects Colln Br. Mus., 13: 1229.
Othreis
fullonia Clerck; Holloway, 1976: 36.

|

|
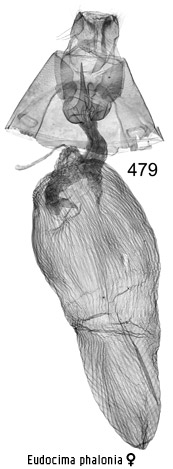
Eudocima
phalonia 
|
Eudocima
phalonia 
|
 |
 |
Diagnosis.
This and the next species have similar sexual dimorphism, the males with more
regularly lineated forewings and the females with more irregularly marked and
mottled ones. In the male of fullonia the fasciation is weak, the oblique postmedial
concave towards the tornus. The female has the postmedial more irregular,
broadly whitened over its posterior half up to a white triangle at the centre;
the reniform is seen clearly darker in a pale surround and has a projection
basad to just posterior to the reniform. See also the next species.
Taxonomic
note. Zilli & Hogenes (2002) reinstated phalonia
as
a senior synonym of fullonia against the advice of Mikkola & Honey (1993), who
indicated that the matter should be referred to the International Commission of
Zoological Nomenclature to maintain precedence of the junior synonym. Zilli
& Hogenes pointed out the economic importance of the species. They noted
that phalonia has been used as valid senior synonym at
least twice since 1899 and that fullonia might
well be a misspelling of phalonia by Clerck. On these grounds they considered that the
requirements for reversal of precedence were not met and that phalonia
should
stand as senior synonym �in the interest of stability of nomenclature�. This
disregards the fact that, in the economic literature, the name fullonia
has
been used for this species, probably exclusively, for at least the last six
decades as indicated by the sole usage of fullonia recorded
by Zhang (1994) in mainly economic literature abstracted on at least 27
occasions over that period. Applied entomologists would therefore probably
disagree with the assertion that reinstatement of phalonia is
in the interests of stability and would prefer to follow the principle advocated
by Sommerer (2002) for the gender issue in nomenclature, that the spirit of the
Code with regard to stability should prevail over the letter of individual
articles when a conflict of interest occurs. Therefore the arrangement of Zilli
& Hogenes is followed with reluctance here.
Zilli
& Hogenes (2002) noted minor but consistent differences in forewing shape
and in the male genitalia of African versus Indo-Australian material of this
species, and suggested that two taxa might be involved.
Geographical
range. Old World tropics and subtropics east to the Society Is.; often
recorded as a migrant, e.g. in Norfolk I. and New Zealand.
Habitat
preference. Frequently encountered from the lowlands up to high altitudes,
the species can be common on mountains where it may have become concentrated by
hill-topping behaviour, e.g. the 56 specimens recorded by Holloway (1976) at
2600m (Radio Sabah) on G. Kinabalu.
Biology.
There are numerous accounts of the life history of this species, e.g. Moore
(1881), Gardner (1941, 1947), Sevastopulo (1941a, 1948), Comstock (1966),
Cochereau (1974), Robinson (1975), Maddison (1982), Tanahara & Tanahara
(2000) and Bell (MS).
The eggs
are laid in loose batches on the undersides of young leaves. The hatchlings are
white, becoming green with lateral brown patches; the setae arise from black
dots. Subsequent instars are black, with orange spots occurring in the position
of the ocellate marks; these do not develop until the fourth instar, then being
rufous, ringed white, sometimes with yellow.
Larger
larvae are pinkish, brownish or rufous, with two diffusely darker longitudinal
bands on each side crossing the dorsal and ventral edges of the ocellate marks
(photograph in Maddison (1982)). The body is irregularly scattered with rufous,
lilac and white specks, the white ones coalescing to form an oblique line up
from the saddle at A6 across A7 as in species mentioned earlier. The ocellate
marks can have an oblique pale dash in the central part at one third from the
ventral edge, and the pale ring is itself surrounded finely by black. There may
be a smaller, solid white mark in the same position on A1. Tanahara &
Tanahara (2000) illustrated a much blacker larva on Okinawa than this, with
darker, diffuse zig-zags laterally. Moore (1881) illustrated a larva with red
patches in the dorsal concavities of the zig-zags and in dorsal bands, but the
manuscript descriptions of other authors that he reproduced are more as in the
first description; the darker bands in the illustration in Maddison (1982) are
more or less straight, and Comstock (1966) illustrated a black larva like that
in Okinawa but more uniform all over. The setae may arise from pale blue spots.
There is
also a green form of the larva. Bell and Sevastopulo referred also to a
grass-green ground colour in India, with the dorsum more pale grey with a
faintly darker dorsal line. There is a subspiracular band of white blotches and
dots, and an oblique extension of this over the saddle by the prolegs as
mentioned above and for other species, and there are white marks associated with
the side of the tumidity on A8. The ocellate marks are bordered thinly with
black then ringed dorsally with cream and ventrally with yellow, but centred by
the green ground colour. The centre of these ocelli may be lined blue or filled
with this colour (Robinson, 1975; Maddison, 1982), and the dorsum may be more
bluish grey, so there is further variability on top of the broad separation into
brown and green forms. The spiracles in the green form are bright crimson.
Host
plants recorded (Moore, 1881; Robinson, 1975; B�nziger, 1982; Tanahara &
Tanahara, 2000; Robinson et al., 2001) are: Leschenaultia
(Goodeniaceae);
Erythrina
(Leguminosae);
Anamirta,
Arcangelisia,
Cissampelos, Cocculus, Coscinium, Cyclea,
Diploclisia,
Legnephora,
Sinomedium,
Stephania,
Tiliacora,
Tinomiscium,
Tinospora (Menispermaceae); Theobroma (Sterculiaceae).
B�nziger (1987) commented on host plant selectivity in Thailand and elsewhere,
indicating that Erythrina is only favoured in the eastern part of the range.
<<Back
>>Forward <<Return to Content Page
|