|
The classification of the Plusiinae, particularly the assignment of
species to genera, is at present far from stable (e.g. the papers of Dufay;
McDunnough, 1942; Eichlin & Cunningham, 1978; Ichinose, 1962, 1973; Chou
& Lu, 1979). It is a problem at present being studied by Dr I.J. Kitching at
the BMNH and so modifications to the arrangement in the systematic account here
are likely in the future. Several species have been included under Plusia
sensu lato because a more precise assignment is not possible until a full
morphological survey of the subfamily has been undertaken.
SUBFAMILY CHARACTERISTICS
Adults
The subfamily is almost certainly a natural group as it can be defined
by numerous characters not found elsewhere in the Noctuidae.
The forewings are triangular with a characteristic hook or falcation to
the dorsum at the tornus. The forewing markings usually include a central pale,
often metallic-looking maculation, either a streak or a ‘u’ and dot that can
be fused to a ‘y’, sited just posterior to the cell. The ‘u’ appears to
be an extension of the orbicular stigma and often grades into the antemedial.
There is often a small fleck of similar pale, bright scales at the posterior end
of the reniform. The venation is generally as in Fig. 15.
The thorax and anterior of the abdomen have a dorsal crest of tufts of
scales that are of a similar colour to the forewings. The males usually have
lateral tufts of long scales on abdominal segments 5 and 6 and sometimes on
others; these are lacking in some of the smaller species in Borneo. The
counter-tympanal hood has a characteristic double structure as seen in Fig. 312
(Richards 1932). The male eighth abdominal segment is modified, usually to a
high degree, but most species have a pair of eversible coremata incorporated in
the sternite as in the two subfamilies already dealt with.

The male genitalia have several unique features. The overall build is
slender, the valves elongate with marginal setae, sometimes a corona, and
usually with a simple, slender harpe at about one third centrally and a clavus
basally, lying adjacent to the juxta. The anellus is finely scobinate in most
species. The saccus often bears small triangular flaps centrally, ventrally. The
aedeagus has the sclerotisation of the shaft restricted to a ventral band and,
except for the nigriluna Walker group, all Bornean species have the base
bulbous with the ductus ejaculatorius inserted distal to the bulbosity. The
vesica is usually extremely long, tubular, invested with rows of large, slender
cornuti, though these are often reduced to a single apical one. There may be
some correlation between the length of the vesica and the bulbosity of the basal
part of the aedeagus: the vesica is relatively short and lobed in nigriluna and
Plusiopalpa adrasta Felder, which have minimal bulbosity, and extremely
long in ‘Plusia’ megaloba Hampson where the bulbosity is extreme.
There is a disc-like or tubular structure basally in the vesica that extends
down the aedeagus shaft.
No constant features were observed in the female genitalia of Bornean
species and their allies that might serve to define the subfamily. The bursa
copulatrix usually lacks a well defined signum but is often generally scobinate.
The antennae of both sexes are filiform in all Oriental taxa.
Larvae
The larvae of Oriental Plusiinae have been studied by Gardner (1947) and
Ichinose (1962, 1973). All those of Bornean species are likely to be of
‘semi-looper’ type with the first and second pairs of abdominal prolegs
absent (small ones are sometimes present in early instars). The crochets on the
prolegs are biordinal (two distinct sizes alternating) and the body setae are
set on distinct pimples. The skin has a covering of points.
Ichinose (1962) divided Japanese species into six groups on the basis of
larval chaetotaxy, but his generic assignations were rather unconventional (e.g.
in Chrysodeixis Hubner); in his later paper (1973) these were more in
line with other treatments in the literature. He drew attention to similarities
in larval characters between Chrysodeixis and Argyrogramma Hubner,
the latter including Ctenoplusia Dufay and Trichoplusia McDunnough
as subgenera. The larvae are characterised by fine setal points. They are
generally green with a very pale line just below the spiracles, and on each side
dorsolaterally there is a broad, slightly paler band flanked by two finer ones.
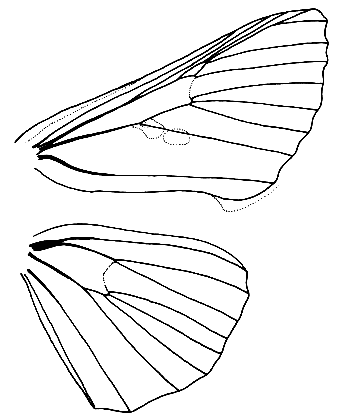
Figure 15. Venation of Chrysodeixis eriosoma, showing
‘u’ and dot maculation and the augmentation of the tornal falcation by the
marginal cilia on the forewing.
Chrysodeixis eriosoma Doubleday and C. acuta Walker larvae occur in
two forms, with and without black dots, these dots usually consisting of two on
each side per segment subdorsally, arranged obliquely, and further subspiracular
spots.
Thysanoplusia Ichinose larvae have different chaetotaxy, and the skin points are large
and hairlike. Though this genus was regarded as distinct from the Chrysodeixis/Argyrogramma
complex by Ichinose it has been associated with Trichoplusia in Inoue
et al. (1982). The former arrangement will be followed here.
Pupae
The pupae of Japanese Plusiinae were described by Nakamura (1974). He
found their characters homogenous, with peculiarities such as the upper position
of the labrum and the rounded projection of the wings and maxillae over the
abdomen that may be diagnostic for the subfamily, though the latter character is
also seen in some Cuculliinae.
Nakamura described the cremaster as ‘robust, fist-like or somewhat
cylindrical and rugose [fluted, grading apically to reticulate, surface
ornamentation], with a pair of long hooked and apically anchor-like setae at
distal end and three pairs of shorter curled setae at dorso-lateral side’, an
arrangement that may also be definitive for the subfamily.
Pupation is in a loose silk cocoon in leaves.
HOST-PLANT RELATIONSHIPS AND ECONOMIC IMPORTANCE
The majority of species are herbaceous feeders; none are recorded from
forest trees in Browne (1968), though CIE records show Argyrogramma signata Fabricius
to feed on Eucalyptus in New Guinea. The following notes have been
collated from Sevastopulo (1941), Mathur (1942), Gardner (1947), Robinson
(1975), Holloway (1977), Brun & Chazeau (1980), and unpublished records kept
by CIE specialists.
Perhaps the majority of species will prove to be polyphagous but
restricted to herbaceous vegetation in disturbed habitats in the tropics, a
category which includes agricultural crops. Several species such as Chrysodeixis
eriosoma and Thysanoplusia orichalcea Fabricius, are recognised pests
and are characteristically widespread geographically, highly dispersive
(recorded as migrants) and usually captured in open habitats.
The only strong indication of specificity is for Anadevidia peponis Fabricius,
widespread in the Indo-Australian tropics but not taken as yet in Borneo. It
appears to be restricted to the family Cucurbitaceae.
Most records of Ctenoplusia albostriata Bremer & Grey are
from the family Compositae. Support for regarding C. vittata Wallengren
(Africa) as sister-species to albostriata is provided by the record by
Pinhey (1975; as transfixa Walker) of a larval host in the Compositae.
Records for other species will be given in the systematic section.
ZOOGEOGRAPHY
The subfamily shows a fairly even distribution of diversity throughout
the world except at very high latitudes.
Just over half the Bornean species are found throughout the Indo-Australian tropics, some extending to the African tropics or having close
relatives there.; Many also extend into the subtropics and migrate into
temperate zones. These widespread taxa are: Chrysodeixis eriosoma, C.
illuminata Robinson, Argyrogramma signata, Thysanoplusia orichalcea,
Ctenoplusia albostriata, Plusiopalpa adrasta, Zonoplusia ochreata, ‘Plusia’
lectula Walker and Dactyloplusia impulsa Walker. The last three
appear to be rare in Australasia.
‘Plusia’ nigriluna is widespread in the Oriental tropics and is replaced by a
sister-species or complex in the Australasian tropics. The minutus Dufay
group of Chrysodeixis includes an allopatric array of species extending
through the Indo-Australian tropics to New Guinea, but the diehli Dufay/ plesiostes
Dufay pair is more or less restricted to Sundaland. Another widespread,
largely allopatric complex of species is seen in the Ctenoplusia tarassota Hampson/sigillata
Dufay group of species, which also extends to New Guinea.
Thysanoplusia bipartita Snellen is shared between Borneo and Sulawesi and
fills a geographical gap between T. intermixta Warren and T. ekeikei Bethune-Baker
but is sufficiently different from those two to cast doubts on the hypothesis
that the three taxa are vicariant products of a widely distributed ancestral
species.
>>Forward
<<Return to Contents page
|