|
The Limacodidae are a moderately diverse, essentially tropical and
subtropical family. There are 95 species recorded from Borneo, 26 in Japan
(Inoue et al. 1982) and only three in Europe. The described world
fauna is in the region of 1000 species though the discovery of numerous new taxa
and complexes in the course of this work would suggest the total to be much
higher.
Their place in the higher classification of the Lepidoptera has yet to
be ascertained satisfactorily. They were placed in the Cossoidea by Brock (1971)
on the grounds of thoracic structure, but Common (1975) included them in the
Zygaenoidea on the basis of early stage characteristics. Fletcher & Nye
(1982) followed Brock's arrangement.
The Megalopygidae, a Neotropical family, are somewhat similar in
external appearance but differ in wing venation as set out in the introductory
key. The larvae are often extremely pilose. The male genitalia lack a gnathus
and typically have setose socii on the tegumen. The females have the bursa
small, flat, opposed ovipositor lobes and a mass of dense, fine pilosity on an
expanded membrane between segments 7 and 8. However, both Fracker (1915) and
Mosher (1916), discussing larvae and pupae respectively, point to shared
characters that could be considered synapomorphic.
Heppner (1984a) placed the Megalopygidae in the Zygaenoidea
(Section Tineina of Ditrysia) and the Limacodidae in the Cossoidea (Section
Cossina of Ditrysia). The justification for this wide separation of the two
families by Heppner is still in press and therefore not available for discussion
here.
Resolution of this uncertainty is beyond the scope of this work but some
pointers are commented on later. It has become apparent, however, from
the Oriental fauna at least, that the family is monophyletic, with
characteristics of both larvae and adults that can be considered apomorphic.
Certain groupings within the family have become apparent, defined by
characteristics of male and female genitalia and venation. These groupings have
led to extensive revision of the systematic arrangement set out by Hering
(1931), and redefinition of the genera has resulted in considerable
recombination of the species.
The family contains numerous species of economic importance. They attack
a wide variety of crops, mainly trees and shrubs such as coconut, oil palm,
coffee, tea and cocoa. It was therefore disturbing to discover that supposedly 'well
known' pest names such as Parasa lepida Cramer, Darna trima Moore,
Thosea sinensis Walker and Setora nitens Walker in fact embraced
complexes of several species. These will be reviewed more extensively than is
possible here in Cock, Godfray & Holloway (in press). Less familiar taxa
also proved on examination more often than not to be more complicated
taxonomically than the literature might indicate.
The Limacodidae are perhaps best known for their slug-like larvae,
sometimes smooth, but usually invested with urticating spines that can cause
profound irritation to the skin, leading to their common name of
�nettle-grubs'.
The larvae of most species appear to be polyphagous, feeding on a wide
range of plant families. No obvious examples of specificity have been located.
Plant families utilised as hosts by species referred to in the systematic
account are ranked as follows by summed records of genera per moth species:
Palmae (25); Rubiaceae, Leguminosae (14); Euphorbiaceae (13); Anacardiaceae
(12); Musaceae (10); Myrtaceae, Theaceae (9); Sterculiaceae (8); Sapindaceae
(7); Combretaceae, Rutaceae (6); Barringtoniaceae (5); Annonaceae, Bombacaceae,
Lauraceae, Lythraceae (3); Dipterocarpaceae, Rhamnaceae, Zingiberaceae (2);
Araceae, Bignoniaceae, Caricaceae, Compositae, Ebenaceae, Ehretiaceae,
Flacourtiaceae, Gramineae, Juglandaceae, Meliaceae, Menispermaceae, Moraceae,
Piperaceae, Pittosporaceae, Punicaceae, Rhizophoraceae, Rosaceae, Salicaceae,
Solanaceae, Stilaginaceae (1).
These records are undoubtedly biassed towards families with genera of
economic importance as records are mainly from literature on pests of crops such
as oil and coconut palm (Elaeis, Cocos), banana (Musa, Musaceae),
coffee (Coffea, Rubiaceae), tea (Camellia, Theaceae), cocoa (Theobroma,
Sterculiaceae), Citrus various (Rutaceae), mango (Mangifera, Anacardiaceae),
and rambutan (Nephelium, Sapindaceae).
Adults
The relatively stout body is usually invested with a dense pile of scales, and
the forewings are also coarsely scaled, the scales arranged haphazardly rather
than in rows.
The antennae are bipectinate in the males of most species, exceptions
being the Narosa Walker group of genera and Trichogyia Hampson and
allies, both falling within larger groupings defined on venation characters
within which this antennal character could define a further subdivision, with
the assumption it evolved independently within each lineage. In most cases broad
bipectination only extends over the basal half to two thirds. The Darna Walker
complex has bipectination to the apex without exception.
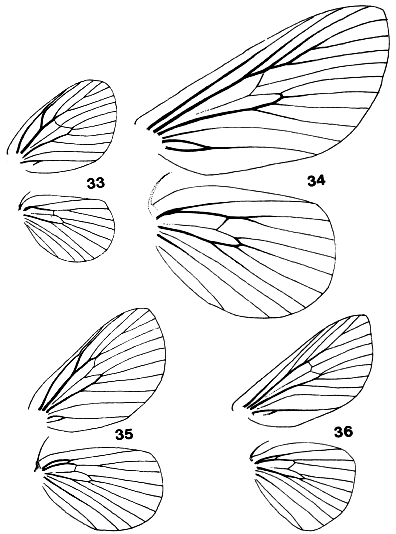
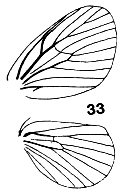
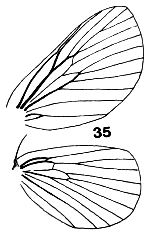
Figures 33-36. Wing venation of Limacodidae. 33, Narosa
concinna; 34, Scopelodes albipalpalis; 35, Cania bandura; 36, Demonarosa
mediodorsata.
The forewings are triangular as a rule, with the stem of vein M
bisecting the cell on both fore- and hindwings. Some wing venations are
illustrated in Figs. 9, 10, 33-39. Figs. 9, 10, 34, 36 are taken to have typical
limacodid ground plan venation, with modifications to the course of R1 in
Figs. 33 & 35, and of the cubital and anal veins in Fig. 38 being taken as
apomorphies to define genera or groups of genera. The stem of vein M is
bifurcate within the cell in Parasa Moore, Hyphorrna Walker, Susica
Walker and Mambarona Hering, but this has not been observed in other
genera.
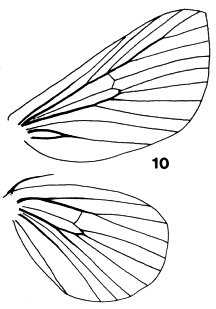 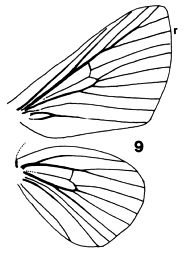
Figures 9-10. Venation
of Limacodidae. 9, Setora cupreiplaga, 10, Setothosea asigna
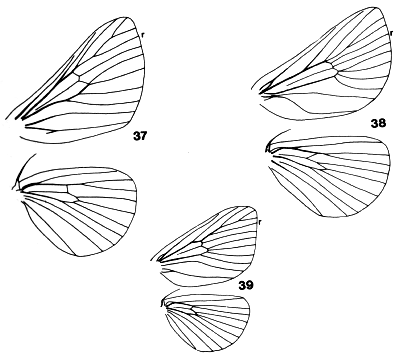
Figures 37-39. Wing venation of Limacodidae showing position of R5 (r)
on the forewing. 37. Nirmides bassalis; 38, Darna metaleuca; 39, Pseudidonauton
admirabile.
The male genitalia are most usually of the simple limacodid ground plan
type as seen in Figs. 134, 153, 161, with a simple elongate valve, a tubular
aedaegus, usually flexed at one third from the base and often with the ductus
ejaculatoris inserting ventrally; scobination or cornuti in the vesica are
unusual, being most widespread in Narosa, Cania Walker and allies. The
uncus and gnathus are typically strongly sclerotised and equal in size. The
gnathus is sharply angled upwards half way along the lateral stems prior to
their fusion into the usually acute distal portion. The apex of the uncus either
tapers uniformly (Fig. 185) or has a more darkly sclerotised ventral spur
apically or just subapically (Figs. 112, 174). These uncus and gnathus
characters may be definitive for the family.
Modifications can occur to this ground plan, some of the most dramatic
being seen in Cania Walker, Susica Walker, Nirmides Hering,
the Darna group of genera and Pseudidonauton Hering; the valves of
Bornean Narosa of the concinna Swinhoe group are bizarrely and
asymmetrically modified.
The female genitalia show a wide range of form, but the coarsely setose,
often slightly bilobed ovipositor lobes form a disc-like pad that is probably
related to the flattened disc shape of the eggs, and are a relatively consistent
feature through the family. The eighth segment bears two ampullate processes
laterally (Figs. 98, 183). The ductus seminalis usually arises from the centre
of the ductus when this is relatively short but subbassally in those taxa with
the ductus more elongate. The ductus bursae is spiralled in representatives of
all the major generic groupings recognised; in some lineages most genera have a
spiralled ductus. When spiralled, the ductus is usually considerably lengthened
and is seen at its most extreme in Phocoderma Kollar where it is
approximately three times the length of the abdomen when extended. The spiral is
brought about by differential sclerotisation in a band running the length of the
ductus, and may represent a definitive feature for the family. The bursa is
usually signate, especially in genera with a spiralled ductus, and the nature of
the signum is used later to define further generic groupings.
|

|

|
The basal stemite of the abdomen is as in Figs. 48, 49 and is of
relatively constant form throughout the family.
The adults have a characteristic resting posture with the body held
almost at right angles to the substrate, propped up on the abdomen, extended
legs and downfolded wings. The posture is illustrated in below and in
Kalshoven (1950, 1981). The moth may suspend itself from a twig in this
position, resembling a piece of dead vegetation.
|

'Thosea' obliquistriga in Hong Kong, showing adult resting
posture (M.J. Bascombe)
|
Eggs
The eggs are
scale-like, somewhat flattened, often laid in an irregular mass (Common 1970).
Larvae
The larvae
of limacodids are perhaps best characterised by the absence of prolegs and the
presence of adhesive, sucker-like zones on the abdomen which sometimes secrete a
viscous, slug-like trail on the host-plant foliage. Locomotion has a peristaltic
quality when seen from below on glass (H.C.J. Godfray, pers. comm.). Fracker
(1915) noted that the thoracic legs are minute and the head is small, retracted
beneath the thoracic segments except when extended during feeding. The shape is
slug-like in many cases but several genera in the groups with a paired or
crescentic signum in the female bursa (see below) have the larva more elongate,
deeper than broad. The dorsal surface terminates just below the spiracles in a
slight flange that is usually maintained in close contact with the substrate.
The ground plan larva is here supposed to have two complete rows of
spined scoli or tubercles on each side, one subdorsal (dorsolateral) and the
other lateral, just above the row of spiracles. The lateral row never includes a
full scolus on the first abdominal segment, where the spiracle is displaced
dorsally from the line of the rest; the scolus is sometimes visible as a small
scobinate patch between the spiracle and the scolus of T3. Modifications include
reduction or enlargement of scoli, differential loss, particularly of the
subdorsals, or complete loss as seen in some lineages discussed later. Some
characteristic larvae are illustrated in Limacodidae
larvaes.
Fracker regarded the Megalopygidae larvae as being transitional to those
of the Limacodidae [Cochlidiidae] within the Zygaenoidea, the scoli having
developed from typical zygaenoid verrucae. In the Megalopygidae there are six
pairs of prolegs, on abdominal segments 2-7 and 10; only those on segments 3-6
bear crochets. There is an additional subspiracular row of tubercles. The spines
of the tubercles or scoli also have urticating qualities but many megalopygid
larvae are highly pilose, the hairs being long and extending upwards and
backwards in a sort of crest.
Pupae
Pupation is in a hard, spherical, almost nut-like cocoon that is often anchored
to the substrate by an irregular arrangement of silken threads. The moth emerges
by cutting out a circular lid at one end.
Mosher (1916) found several resemblances to the pupae of Megalopygidae:
it is typically stout; there are similarities in the motility of abdominal
segments; in head sutures; in characteristic eyepieces. The eyepiece, a
sculptured flange, appears to be unique to Limacodidae and Megalopygidae and
could therefore be a synapomorphy for the two families.
Comments on
relationships
The introductory key indicates that some genera currently in the Megalopygidae,
such as Aidos Hubner, have limacodid-like forewing venation and antennae.
The male genitalia and female ovipositor lobes of Aidos have more in
common with those of the limacodids than with those of Megalopyge Hubner
and Norape Walker. The status of these genera and the naturalness and
relationships of the two families needs further investigation; the characters of
early stages are likely to prove important in this.
Brock (1971) suggested that forewing venation in his Cossoidea has
evolved through the 'splitting-back' or migration basad down R5 towards
the cell of vein M1 and the radial veins. This has not occurred in
the Megalopygidae but has for R5 and sometimes R4 (Fig.
38) in the Limacodidae. If this transformation series is correct then it
provides a further apomorphy for the Limacodidae relative to the supposed
sister-group, the Megalopygidae, and would indicate that Aidos and
relatives should be transferred to the Limacodidae. It might then be possible to
test the monophyly of the Megalopygidae with reference to the characters of male
and female genitalia mentioned above.
The evolution of the limacodid larva could then be interpreted as
envisaged by Fracker (1915) in terms of: loss of the subspiracular tubercles and
development of a subspiracular flange; loss of crochets from prolegs on segments
3-6 and extension of adhesive zones into a more or less continuous surface;
retraction of the head under the thoracic segments.
Groupings of genera in the Oriental fauna
The survey of male and female genitalic morphology necessary to establish the
identities of Bornean limacodid species and to define the genera to which they
are assigned has revealed certain characters whose distribution suggested a
classification somewhat at odds with that established by Hering (1931) for the
Oriental fauna. It has not been possible to assess these discoveries in relation
to the African or New World faunas but, given that New World species such as Parasa
chloris Herrich-Schaffer and Miresa clarissa Stoll are obviously
congeneric with Oriental species, it is probable that these groupings will be of wider relevance. Janse (1964) followed Hering
(1955) in relying mainly on venation and leg characters for keying African
genera, though he did examine the genitalia for most species and describe them
for each genus.
The spiralling of the female ductus bursae may well be a limacodid
ground plan character that has been lost independently in most of the lineages
discussed below. It is mentioned for only very few African genera by Janse
(1964), most of which either lack signa in the bursa or are bisignate.
Subdivisions of the Oriental fauna can be made on bursa ornamentation,
though it is unclear which, if any, of the states is plesiomorphic. Here it is
assumed as a working hypothesis that general uniform scobination is
plesiomorphic and that the signate character states observed can be used to
define distinct lineages.
In the first lineage the bursa contains an elongate, somewhat elliptical
field of small, rounded, stellate-spined signa. These are most numerous in
typical Narosa Walker, Belippa Walker and the temperate Oriental
genus Austrapoda Inoue. The number of signa is reduced, with enlargement
of those that remain, in Apoda Haworth (of which Limacodes Berthold,
the type genus of the family, is a synonym) and Demonarosa Matsumura
(Fig. 100). A single round signum is seen in Cania Walker and Altha Walker;
within both these genera there are species where the spiral of the ductus has
been lost. These two genera are linked to Narosa by venation and facies
characters respectively. Signa and spiral are weak or lost in Nagoda Moore
and Cheromettia Moore but these share, in the females, a similar mottled
facies with other taxa. Chalcocelis Hampson and Chalcoscelides Hering
are also tentatively associated with this group on grounds of facies, sexual
dimorphism and larval characters.

Many genera showing strong sexual dimorphism are included in the group: Cania
(part), Chalcocelis, Cheromettia, Belippa, Nagoda. Most have larvae
with the scoli absent (Altha, Narosa, Cheromettia, Chalcocelis,
Chalcoscelides) but in Cania they are merely reduced. Outside the
group, only Birthamoides Hering (crescent-signum group) has the larva
smooth-skinned without scoli.
The genus Atosia Snellen and three new genera (Flavinarosa,
Heringarosa and Saccurosa) are associated with Narosa through
sharing filiform antennae in the male and a sinuous vein R1 in the
forewing that creates a distinctively broad subcostal area (Figs. 33, 35). This
venation character is shown strongly by Cania, Mambarilla Hering, and to
some extent by Altha and Althonarosa Kawada.
 |
 |
In the majority of genera the palps are upcurved with the third segment
conspicuous. The second and third sections are not clearly distinguished from
each other. The crescent-shaped signum of the third section (Fig. 218) may be a
development from the second through fusion and lateral elongation of the paired signa (Figs. 164, 173) of the latter; a possible
intermediate condition, with a pair of scobinate signa compressed across the
bursa and linked by a ridge, is seen in the Indian Iragoides crispa Swinhoe.
The crescent signum group has not been located outside the Old World tropics but
the bisignate condition is found pantropically.
In both sections genera have larvae where the subdorsal rows of scoli
are heterogeneous in size, often extremely so. A common adult forewing pattern
in both groups is of dark submarginal and postmedial fasciae angled to each
other, the latter often passing obliquely across the wing close to the discal
area where there may be a dark discal spot. The palps, with some notable
exceptions (Scopelodes Westwood and allies), are directed forwards,
short, deep, the third segment short and obscure (Fig. 101).
|
|
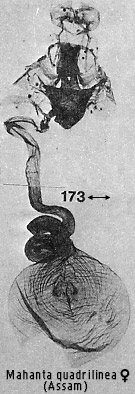
Genera in the paired signa section are: Miresa Walker, Parasa Moore,
Hydroclada Meyrick (Australasian tropics only), Hyphorma Walker, Mahanta
Moore, and Phocoderma Butler. Hyphormides Hering and Scopelodes
form a group with Hyphorma as they share a distinctive elongation of
the palp, particularly of the third segment (Figs. 157-159). Females of Hyphormides
have not been located; Scopelodes females are bisignate but, unlike
the other genera, have the ductus short, unspiralled. Susica Walker is
tentatively associated with this group as its larvae are generally similar, but
the female bursa lacks signa and the ductus is straight.
The crescent signum section includes the genera Thosea Walker, Praesetora
Hering, Setora Walker, Birthamoides Hering, Birthamula Hering,
Allothosea Hering, and the new genera Setothosea, Birth osea and Griseothosea.
Most of these genera have pale patches at the apex of the foretibia, some
having a similar spot at the apex of the forecoxa. Many have a furca from the
base of the sacculus in the male genitalia. The larvae have the anterior and
posterior dorsolateral scoli much larger than those on A2-7 as in the bisignate
group, but that on A5 is also enlarged; the larva of Thosea is
exceptional both in shape and in the uniformity of its rows of scoli.
The genera Birthama Walker and Idonauton Swinhoe are
tentatively associated with these two sections for reasons given in the generic
accounts.
The genera Pseudidonauton Hering, Nirmides Hering and,
both newly described here, Pseudonirmides and Bornethosea cannot
be assigned to any of these groups on adult features at present. The larva of Pseudonirmides
is distinctive. The genera lack the venation modifications described for the
final section.
The final section is defined on modifications of the forewing venation
(Fig. 38). Vein R5 is 'split-back' to arise independently from the
cell at a small distance from Rs. The cubital veins are usually curved to a
lesser or greater degree and the anal vein or CuP can be sinuous. The
splitting-back of R5 is evident in both sexes but the other venation
features tend to be more strongly developed in males.
Figures 37-39. Wing venation of Limacodidae showing position of R5 (r) on
the fore-wing. 37, Nirmides basalis; 38, Darna metaleuca; 39, Pseudidonauton
admirabile.
Two subgroups can be recognised. One has filiform antennae in the male
and a somewhat divided valve to the genitalia in some genera. It includes Ceratonema
Hampson, Trichogyia Hampson, Caissa Hering, Limacosilla Hering,
and a new genus, Striogyia. Euphlyctinides Hering also belongs to this
subgroup, but the type species, rava Hering, is a junior synonym of Ceratonema
albifusum Hampson, syn. n. The other subgroup has male antennae completely
bipectinate and includes Darna Walker and two new genera, Marsuplectra
and Limantricodes.
The larvae of this final section tend to be somewhat humped towards the
anterior, the dorsolateral scoli much smaller than the laterals which are often
longer posteriorly.
Zoogeography
Of the 95 Bornean species, 35 (37%) are at present only known from
Borneo. Five are shared with Sumatra only and ten with Peninsular Malaysia
only. The Sumatran fauna has not yet been studied in depth but material
of the Heterocera Sumatrana Society will shortly be studied by the author and
may change this picture dramatically as it has already done with the
Notodontidae. Further collecting in the Peninsula is also needed.
28 species are found in at least three of the major land areas of
Sundaland (Peninsular Malaysia, Sumatra, Java, Borneo) but not outside except
perhaps in the south of Burma or Thailand. Thus about 83% of the Bornean fauna
is restricted to Sundaland.
The remainder includes one species shared between Borneo and the N.E.
Himalaya (Flavinarosa holoxanthia Hampson), one between Borneo and
Palawan (Thosea borneensis Hering), one between Borneo and the
Philippines (Pseudonirmides sola Swinhoe) and one between Sundaland and
Sulawesi (Thosea vetusta Walker). Two species extend from Sundaland
eastwards (Mambarona congrua Walker, Chalcocelis albiguttata Snellen)
and the rest are shared between Sundaland and parts of mainland Asia: with China
(Darna pallivitta Moore); with the N.E. Himalaya and sometimes Taiwan (Cania
bandura Moore, Chalcoscelides castaneipars Moore, Praesetora
albitermina Hering, Idonauton apicalis Walker, Trichogyia
semifascia Hampson); with the Indian Subregion (Phocoderma velutina Kollar,
Birthamoides junctura Walker) and with the Oriental tropics generally (Miresa
bracteata Butler, Hyphorma minax Walker).
Three genera, newly described here, are apparently endemic to Borneo (Bornethosea,
Marsuplectra, Saccurosa) and seven more are restricted to Sundaland (Birthosea,
Heringarosa, Hyphormides, Limacosilla, Nirmides, Setothosea, Striogyia). The
rather distinctive lawaii group of Narosa appears to have radiated
extensively in Sundaland and the Philippines, with a more distant relationship
to mainland Asia. Similarly, the genera Atosia, Arbelarosa, Birthamula, and
the Cania bandura group are most diverse in Sundaland with one species in
mainland Asia.
Mahanta consists of a Himalaya + Taiwan/Sunda sister pair and reflects Chalcoscelides
in overall distribution. Two endemic species of Nagodopsis have one
other congener only in Taiwan.
Several Bornean/Sundanian species are outliers of genera or groups that
are most diverse in mainland Asia such as in Cheromettia, Miresa and the Cania
bilinea Walker group.
Most of the remaining genera are diverse throughout the Oriental tropics
and some, such as Scopelodes, Narosa, Chalcocelis and Thosea extend
to the Australasian tropics, the first and last strongly so. Birthamoides consists
of an allopatric array of species that extends from India to the Solomons. The
area relationships of the species reflect general area relationships revealed by
an analysis of butterfly and bird species distributions (Holloway & Jardine
1968; Holloway 1984a) and match those of a number of other allopatric
arrays in the Indo-Australian tropics (Holloway 1982b, in press).
A more complex pattern, perhaps of more ancient establishment, that is
inconsistent with these area relationships is seen in the concinna Swinhoe
group of Narosa, with two Sundanian species sister to a Sulawesi/New
Guinea complex (Holloway in press). Relationships within Thosea also show
the Sulawesi fauna to be allied to that of New Guinea and the Philippines rather
than to that of Borneo (p. 104). In Setora the Sulawesi and Philippines
species form a group distinct from those of Sundaland and most closely allied to
taxa in mainland Asia. Both Thosea and Setora will be discussed in
more detail in Cock, Godfray & Holloway (in press). Direct links between
Sulawesi and Borneo are seen at a specific level only (Chalcocelis
albiguttata, Thosea vetusta, Mambarona congrua). The relationship between
Borneo and Sulawesi is thus a distant one despite current geographical
proximity. This apparent paradox will be discussed in relation to the geological
history of Sulawesi elsewhere (Holloway, in press).
|
|
A recurring pattern of area relationships is evident within Sundanian
Limacodidae, illustrated by the general area cladogram in Fig. 40. The cladogram
is fully dichotomous (i.e. fully resolved) but some species groups, whilst
consistent with it, show less resolution, variations on the theme, or only part
of the pattern.
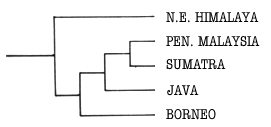
Figure 40. Cladogram showing general area relationships shared by groups of
Limacodidae as discussed in the text.
In most instances complete allopatry is not evident and one can only 'retrieve' this general pattern by making the assumptions suggested by
Holloway (1982b) that speciation is always allopatric and dispersal
parsimonious. These assumptions can generate similar area cladograms to those
derived by applying the method of Nelson & Platnick (1981: 447-9, assumption
2) for dealing with widespread taxa and redundant distributions, but this is not
always the case.
In the cladogram for Darna (Fig. 41, below), the subgenus Ploneta
is consistent if one assumes that diducta dispersed to Borneo to
overlap with bradleyi; Sumbawa is included and the Himalayan outgroup is
multiple.

Figure 41.Cladogram
for the genus Darna indicating the subgenera recognised. Numbers instead of
species at the branch ends refer to BM genitalia slide numbers of specimens
belonging to undescribed species. The groupings within the cladogram are defined
by apomorphies listed in the text under the numbers on the diagram.
Distributions of the species are indicated by the following abbreviations:
B.Borneo; C, China; IN, India; J.Java; M, Peninsular Malaysia; N.E. Himalaya;
N.IN, North India; PH, Philippines; SL, Sri Lanka; S.IN, South India; SUL,
Sulawesi; SUMB, Sumbawa; SUN, Sundaland; TH, Thailand.
Within subgenus Orthocraspeda the sordida pair is
consistent with the Sundanian part of the general area cladogram but there is no
resolution of the Peninsula/Sumatra/Java triplet. Complete resolution is shown
by the trima complex if it is assumed that trima ajavana has
subsequently spread to Borneo.
Another example is seen in Atosia where a subsequent dispersal of
doenia to Borneo must be assumed, and where the Bornean branch is
represented by a species pair. Again the Peninsula/Sumatra/Java triplet is
unresolved. In the more complex scenario for Susica outlined by Holloway
(1982b) the malayana and pannosa groups both show
resolution of Sundanian areas, though the former is not represented in Java, and
the sister group of both lineages is suggested to be Himalayan by the allopatry
+ parsimony approach.
Thosea loesa + mediostrigata as sister pair to T. borneensis is also
consistent, though material from Sumatra is lacking.
Thus examples of this pattern total seven. Counter examples are fewer,
such as Scopelodes testacea (Himalaya) + anthela (Peninsula and
Sumatra) versus albipalpalis (Borneo), and perhaps the Cania bandura group
when the relationships of the taxa have been resolved.
The pattern is also counter to the relationship between the major areas
of Sundaland suggested by the phenetic analysis of overall faunistic similarity
described by Holloway & Jardine (1968). In analyses for both birds and
butterflies Borneo is sister area to the Peninsula + Sumatra, with Java the
outlier. However, this disagreement may be due to the frequency with which the
Peninsula + Sumatra taxon in the pattern has broken allopatry by dispersal to
Borneo, increasing the faunal similarities of the three areas.
Such consistent area patterns may indicate something of the sequence of
barrier development relative to Limacodidae in South East Asia, following a
period of ecological uniformity or general dispersal, perhaps during Pleistocene
climatic fluctuations. Fixation of such events in time or in relation to other
evidence of geographic changes will be exceedingly difficult. In addition, as
suggested by the Susica example for instance, the interchange of periods
of uniformity/dispersal with periods of physiogeographic fragmentation may have
been cyclic (Holloway 1973, 1982b).
Habitat preference
In Borneo the majority of species are found predominantly in the lowlands. Only
about 14 are known only from lower or upper montane habitats and many of these
are represented by only one or two specimens.
During the Mulu survey the greatest limacodid diversity was encountered
in the wet heath (Kerangas) forest at about 150m on sandy raised terraces by the
S. Melinau (Holloway 1984b). Collections from Brunei produced a number
of interesting limacodid records from drier heath forest, swamp forest and mangrove. The family may therefore be relatively more
frequent than are other Lepidoptera groups in lowland forest types on acid
soils, though more observations are needed.
>>Forward
<<Return to Contents page
|